Новый сезон культового сериала «Ольга» возвращается на ТНТ — Пермский информационный портал — 59i.ru
Самый народный комедийный сериал страны возвращается после двухлетнего перерыва в эфир канала с четвертым сезоном.
В новом сезоне Ольга дистанцируется от семьи, и та оказывается на грани катастрофы. Как описывает ситуацию неудавшийся жених Ольги Гриша: «Аня жениха выгнала, Тимофей со шпаной связался, дядя Юра вообще цементом могилы заливает… а ей фиолетово!»
Только возвращение матери-героини может спасти семью… Но захочет ли Ольга в очередной раз быть самой сильной и решать за всех?
Герои «Ольги» вовсе не пропали с радаров — они взяли паузу в два года, чтобы аккумулировать силы, заметно подрасти (касается молодых актеров) и дать бой суровой жизни и здравому смыслу, используя фирменный арсенал суперспособностей: умение вляпаться без каких-либо предпосылок к этому, сломать ногу на ровном месте, устроить пожар в бане или потоп в бассейне. Дирижирует оркестром косячных по-прежнему Ольга (Яна Троянова).
Дирижирует оркестром косячных по-прежнему Ольга (Яна Троянова).
В новых сериях комедийного проекта ТНТ семейство Терентьевых начинает безудержно спасать окружающих. Чего бы им и спасаемым это не стоило. Сестра Ленка (Алина Алексеева) вызволяет молодого отца своего ребенка Пушкина (Филипп Ершов) из армейской части и готовится к свадьбе, дочь Аня (Ксения Суркова) помогает странному Льву (Юрий Борисов) научиться заботиться не только об окружающих, но и о ней с их ребенком, сын Тимофей (Мухаммед Абу-Ризик) спасает соседку Сашу (Анастасия Матвеева) от неблагополучного отца, бухарика Жоры (Павел Майков), параллельно сам впутываясь все глубже в криминальные истории. Даже Юрген (Василий Кортуков) хочет помочь жене, которая уже умерла.
А что Ольга? Впервые за четыре сезона героиня пришла к тому, чего требовала логика, организм и драматургия — надломилась и отпустила ситуацию в семье, решив — впервые за все сезоны сериала — заняться собой.
Яна Троянова: «Вообще, это иллюзия, что у Ольги в руках вожжи.
Удастся ли Ольге справиться с первым серьезным кризисом в жизни и принять реальность такой, какая она есть? Что случится со взрослыми детьми, привыкшими к гиперопеке со стороны матери? Как проявятся в этой ситуации несостоявшиеся мужчины Ольги — шеф гробовщиков Гриша (Максим Костромыкин), добрый бизнесмен Володя (Гоша Куценко) и вредный следователь (Олег Тополянский)? И нужна ли она вообще кому-то, кроме себя?
Смотрите новый сезон комедийного сериала «Ольга»(16+) с 31 августа в 20. 00 на ТНТ!
00 на ТНТ!
На ТНТ начинается новый сезон популярного сериала «Ольга»
Она по-прежнему по утрам спешит на работу в салон красоты, на очередной сеанс педикюра. После работы бежит в сетевой супермаркет, чтобы закупиться товарами по акции и принести добычу в дом, где ее ждет большая, голодная и непутевая семья.
Во-первых, двое детей от двух разных браков, которых она продолжает воспитывать в привычной манере «Я консерватории не заканчивала, но если чего — пюпитр оторву».
Во-вторых, отец, пенсионер-бездельник-алкоголик, бывший футболист, соблюдающий режим завязки-развязки: «Уже третьи сутки держусь, даже фломастеры не нюхал». А с некоторых пор в их доме поселился Гриша. Простой рязанский парень, работает водителем катафалка и по собственному заявлению — будущий муж Ольги.
Еще в квартире ошивается парень дочери Андрей и сестра Ольги Лена. От этих двух персонажей тоже никакой помощи, сплошные убытки, поэтому за ними постоянно приходится приглядывать.
Вот такой неподъемный груз ежедневно ложится на хрупкие плечи Ольги. Такое по силе только настоящей русской женщине, матери-героине, способной останавливать на скаку коней прямо посреди горящей избы.
В новом сезоне ситкома «Ольга»:- Старшая дочь вот-вот родит, а ее парень загремит в полицию за мошенничество. Дело передадут старшему следователю, который положил свой полицейский глаз на Ольгу и уже придумал как решить проблему.
- Отец Ольги больше не пропивает последние деньги, окружающие, в его присутствии, не дышат перегаром, а сам он всегда в отличном настроении. Правда при этом он получит новую необычную зависимость.
- Гриша со своей бабушкой постоянно влезают в дела Ольги и учат жизни. Молодой (все-таки младше Ольги на 12 лет) человек уже не просто регулярно подкатывает к подъезду на своем катафалке, а застолбил постоянное место на парковке возле ее дома.
 Он собирается жениться на Ольге и завести детей. Только Ольгу он об этом забыл спросить.
Он собирается жениться на Ольге и завести детей. Только Ольгу он об этом забыл спросить.
 Он собирается жениться на Ольге и завести детей. Только Ольгу он об этом забыл спросить.
Он собирается жениться на Ольге и завести детей. Только Ольгу он об этом забыл спросить.Проблем много, но Ольга обязательно со всем справится. Потому что других вариантов нет.
«Если честно, то волнение, как будто у нас стартует не второй сезон, а премьера, — говорит Яна Троянова, исполнительница главной роли в сериале „Ольга“. — Хотя при этом есть уверенность в том, что новый сезон получился сильнее. Во всяком случае по сюжету он точно глубже. Линия моей героини стала более драматичной. Но как бы не складывались события, Ольга всегда должна быть сильной и делать выбор в пользу семьи. И пусть ради этого ей даже приходится переступать через себя».
Фото: пресс-служба ТНТ.
Ольга (Ольга) — Сериалы — смотреть онлайн легально на MEGOGO.RU
Этот сериал будет доступен на MEGOGO до 31 декабря 2018 года.
Жизнь прожить — не поле перейти, у каждого человека существует своя жизненная история. У кого-то путь по жизни был, возможно, не сильно трудным, ну а кто-то всё время пытается карабкаться по своей лестнице вверх и у него это не очень ловко получается. Всегда могут быть на дороге различные препятствия, недомогание и другие причины, способствующие этому помешать.
Всегда могут быть на дороге различные препятствия, недомогание и другие причины, способствующие этому помешать.
Так и фильм Ольга рассказывает о повседневной жизни простой русской матери, которую зовут Ольгой. Эта женщина является любящей мамой своих двух уже немаленьких детей: старшей дочери Анны, которой уже исполнилось шестнадцать лет и младшего сына одиннадцати лет Тимофея. Они уже достаточно долго живут в квартире из двух комнат, кроме них здесь ещё проживает и отец Ольги Юрий, бывший футболист на пенсии, да и ещё не прочь хорошенько выпить и родная сестра Лена. Оля всё своё время уделяет семье, заботится о детях, которым ни капельки не хотят помогать их отцы, о своём отце-пьянице, о домашнем уюте. За что ни разу, ни от одного домочадца не услышала ни единого комплимента, благодарности, а только постоянные укоры да нарекания. Квартира Ольги расположена в одном тихом районе города Москвы. Сама же она работает парикмахером в не очень дорогом салоне и всячески старается выкручиваться, чтобы содержать детишек.
Аня, которая постарше, вовсе не подчиняется просьбам матери, считает себя достаточно взрослой и самостоятельной, старается жить, как ей кажется, очень правильно. Но увы, это только создаёт ещё больше проблем для матери, которая и так уже сбилась с ног, решая всё за всех. Сын Тимофей, которому едва перевалило за первый десяток — хочет всем показать, что он настоящий уже мужчина. Ну а с сестрой всё предельно ясно — это ленивое создание существует при помощи чужих мужей. Однако наша героиня по жизни оптимистка и никогда не покажет на улице свою усталость или растерянность, она весело будет шагать дальше, преодолевая все трудности.
Этот сериал будет доступен на MEGOGO до 31 декабря 2018 года. Жизнь прожить — не поле перейти, у каждого человека существует своя жизненная история. У кого-то путь по жизни был, возможно, не сильно трудным, ну а кто-то всё время пытается карабкаться по своей лестнице вверх и у него это не очень ловко получается. Всегда могут быть на дороге различные препятствия, недомогание и другие причины, спосо
Феномен сильной российской женщины в сериале «Ольга» с Яной Трояновой
На телеканале ТНТ после двухлетнего перерыва выходит новый сезон сериала «Ольга» — истории о матери-одиночке, которая на своих хрупких плечах тянет весь мир.
В обычном московском районе Чертаново живет семья Терентьевых — глава семейства Ольга, ее пьющий отец, бывший физрук ЮрГен, дочка Аня, которая думает, что она умнее всех и точно выберется из тесной панельки, сын Тимоша — ему всего 11, но он уверен, что он уже настоящий мужчина, и сестра Ленка, зависимая от мужчин. На горизонте у Ольги периодически появляются разные ухажеры — то трогательный и наивный водитель катафалка, то крепкий и решительный бизнесмен, но ни один из них не остается с Ольгой надолго, потому что, по словам актрисы Яны Трояновой, не может вынести рядом с собой такую сильную женщину, которая рвется решать все проблемы сама.
Вообще, что значит «сильная женщина» в массовом кинематографе? Как правило, это бизнесвумен — успешная в работе, несчастная в личной жизни, это героиня, которой пришлось столкнуться с болезнью, это героиня, которой пришлось пройти через потерю близких,— и всех их в финале ждет новая счастливая любовь. Именно такой финал наши кинематографисты считают наградой сильным женщинам за их страдания, как будто и не задумываясь о том, что давать надежду, поддержку и веру в жизнь одиноким женщинам можно совершенно иначе, без искусственно созданных отягчающих обстоятельств.
Можно, как создатели «Ольги», придумать для героини не катастрофы планетарного масштаба, чтобы показать ее силу, а рутинные проблемы вроде сорванной поездки в Турцию, ремонтного бедлама, разборок то с гулящей дочкой, то с гиперактивным сыном, усталости от работы и вечного желания помочь всем. И зрителям это понравится, потому что это на самом деле про них — простых людей, которым хватает заморочек в неустойчивой российской реальности. В этом и есть феномен сильной женщины Ольги, чьи проблемы знакомы каждой. «“Ольга” вообще очень актуальна и своевременна — как разговор о женщинах-одиночках, разведенках. Это вера для женщин в то, что мы прорвемся. Ведь им тяжело живется одним, какими бы сильными они сегодня ни были. Как бы они ни скрывали свою боль — им больно. А когда такие персонажи, как мой, приходят к ним в дом — они их поддерживают, и женщины начинают в себя верить. Ольга говорит: “Верь в себя!”» — считает Яна Троянова. «Нам показалось, что это очень по-русски, когда есть женщина, на которой все держится»,— говорит продюсер сериала Артем Логинов.
В этом и есть феномен сильной женщины Ольги, чьи проблемы знакомы каждой. «“Ольга” вообще очень актуальна и своевременна — как разговор о женщинах-одиночках, разведенках. Это вера для женщин в то, что мы прорвемся. Ведь им тяжело живется одним, какими бы сильными они сегодня ни были. Как бы они ни скрывали свою боль — им больно. А когда такие персонажи, как мой, приходят к ним в дом — они их поддерживают, и женщины начинают в себя верить. Ольга говорит: “Верь в себя!”» — считает Яна Троянова. «Нам показалось, что это очень по-русски, когда есть женщина, на которой все держится»,— говорит продюсер сериала Артем Логинов.
Ассоциируют себя россиянки и с неунывающей женщиной, внутри одинокой, снаружи — окруженной людьми, чьи проблемы надо решать, деловой, заботливой, думающей в первую очередь о других, во вторую — о себе. А самое интересное, что в «Ольге» — несмотря на то что это ситком и по его законам главный герой должен быть примером, на который равняются,— мать двоих детей вовсе не показана как женщина, все делающая правильно. Ольга неидеальна — она вспыльчивая, она часто берет на себя больше, чем может вынести, порой она откровенно забивает на себя и потом пожинает плоды депрессии. Она просто такая, какая есть,— этим и цепляет. И в новом сезоне ее ждут трудности, которые закономерно вытекают из обстоятельств ее жизни.
Ольга неидеальна — она вспыльчивая, она часто берет на себя больше, чем может вынести, порой она откровенно забивает на себя и потом пожинает плоды депрессии. Она просто такая, какая есть,— этим и цепляет. И в новом сезоне ее ждут трудности, которые закономерно вытекают из обстоятельств ее жизни.
«Ольга» — с 31 августа в 20:00 на ТНТ!
16+
Сериал Ольга 3 сезон смотреть онлайн все серии
Вот наконец мы дождались сериала Ольга 4 сезон на ТНТ Премьер (Смотри у меня) — комедийный сериал о простой русскую семью, будни которых и отражены в сериале. Этот сериал полюбился зрителю по своей простоты и раскрытия тяжелой судьбе русской женщины, которая воспитывает малолетних детей сама, и к тому же отец пьет каждый день. Третий сезон будет намного интереснее, и более наполнен комическими эпизодами — не стали бы режиссеры снимать продолжение не дожидаясь премьеры первого сезона, они были уверены на все сто в успешности своего творения. Описание 4 сезона сериала Ольга на ТНТ — первый сезон закончился довольно неоднозначно, осталось очень много вопросов, на которые нет ответа. Хорошо то что Аня не успела сделать аборт, правда из-за этого Андрюха попал за решетку. С помощью подруги Ани удается вытащить его из следственного изолятора, но и здесь есть свои нюансы. Теперь на глазах отца подруга должна ходить с парнем, ведь отец девушки думал что помогает своему будущему зятю. Через эту ситуацию подружки поссорились.
Хорошо то что Аня не успела сделать аборт, правда из-за этого Андрюха попал за решетку. С помощью подруги Ани удается вытащить его из следственного изолятора, но и здесь есть свои нюансы. Теперь на глазах отца подруга должна ходить с парнем, ведь отец девушки думал что помогает своему будущему зятю. Через эту ситуацию подружки поссорились.
Каждая новая серия третьего сезона Ольги гораздо смешнее и интереснее предыдущих, и для того чтобы узнать ее сюжет, можно просто посмотреть анонс, в котором будет рассказано об основных повороты следующего эпизода. Это все бесплатно можно на нашем сайте.
Также практически каждый день появляются новые видео, в которых рассказывается об актерах и их героев, даже в некоторых они сами рассказывают о своих сюжетных персонажах. Их фото, с их личной жизни и сериала можно найти в разделе «Музыка, фото и видео», которые помогут более подробно почувствовать жизнь героев.
В общем Оля во 4 сезоне продолжает решать ежедневные проблема матери одиночки — помогает молодой дочери Ани растить ребенка, она сама по глупости стала матерью в такой же возраст. Сын Тимофей как можно быстрее старается повзрослеть, и стать главой своей семьи, ведь он один мужского пола. Новоиспеченная бабушка продолжает извлекать своего отца из запоя, но он снова обманывает свою спутницу жизни, и по его словам в лечебных целях продолжает бухать. На счет жизни ЮрГен будет выделена отдельная сюжетная линия. Ну а Андрюха узнав о рождении ребенка сразу же повзрослел, и станет хорошим семьянином, конечно же без простых будничных проблем в новой семье не обойдется. Подведя итог, приглашаем всех смотреть в хорошем качестве продолжения любимого российского комедийного сериала на ТНТ, премьера которого состоялась осенью 2020 года.
Сын Тимофей как можно быстрее старается повзрослеть, и стать главой своей семьи, ведь он один мужского пола. Новоиспеченная бабушка продолжает извлекать своего отца из запоя, но он снова обманывает свою спутницу жизни, и по его словам в лечебных целях продолжает бухать. На счет жизни ЮрГен будет выделена отдельная сюжетная линия. Ну а Андрюха узнав о рождении ребенка сразу же повзрослел, и станет хорошим семьянином, конечно же без простых будничных проблем в новой семье не обойдется. Подведя итог, приглашаем всех смотреть в хорошем качестве продолжения любимого российского комедийного сериала на ТНТ, премьера которого состоялась осенью 2020 года.
С самого старта новой серии команда поддержки Ольги бросилась спасать Тима, которого загребли в СИЗО за нападение на инвалида. Каждый предлагает свою версию событий в сериале Ольга 4 сезон 12 серия на ТНТ: Наташа, благодаря своей интимной работе, знает одного влиятельного прокурора, по совместительству являет клиентом, предлагает обратиться к нему. Гриша направляется к следователю, попытается… Читать далее »
Гриша направляется к следователю, попытается… Читать далее »
Увлекательный сериал Ольга 4 сезон 11 серия продолжает удивлять нас своими неожиданными поворота сюжета, а также нестандартные поступками главных героев. Семья Терентьевых совсем не скучает, ведь Тимофей попал в серьёзную беду.
Раздел: Ольга 4 сезон смотреть онлайн до эфира все серииСериал Ольга с сильной героиней продлили на 4 сезон, в котором ключевому персонажу приходится всерьёз разбираться с усложнившейся жизнью собственной семьи. Сериал Ольга 66 серия вновь расскажет нам о том, как личная жизнь героини налаживается, она выпивает и веселится вместе с Володей, а время ненадолго перестаёт для, неё существовать. Юрген продолжает настаивать на том, чтоб Елена повлияла на свою сестру. Теперь им приходится коллективно устраивать авто подставу с неизвестными последствиями.
Теперь им приходится коллективно устраивать авто подставу с неизвестными последствиями.
После всех последних событий жизни Ольги потихоньку начала налаживаться. Дети стали ее лучше понимать и практически перестали создавать проблемы и помогают по хозяйству. В сериале Ольга 57 серия онлайн она наконец может спокойно работать в сало и зарабатывать деньги на семью, а не бегать и решать ее проблемы. Саша увидела в Тимофее настоящего мужика, который может заступиться за свою девушку, и перестала относиться к нему как к малолетнему — они начали официально встречаться.
Раздел: Ольга 4 сезон смотреть онлайн до эфира все серииЗаключительная серия третьего сезона даст ответ на многие вопросы, которые целый сезон интересуют зрителей. Поэтому прямо сейчас начинайте смотреть сериал Ольга 56 серия онлайн бесплатно, и узнайте как отреагирует Оля предложение Гриши, который хочет забрать ее с детьми подальше от города. Главный вопрос сезона — поедет Оля с Гришей и оставит все свои заботы на старом месте жительства
Главный вопрос сезона — поедет Оля с Гришей и оставит все свои заботы на старом месте жительства
Канделаки стала исполняющим обязанности директора ТНТ — РБК
Тина Канделаки (Фото: Вадим Тараканов / Global Look Press)
«Газпром-Медиа» назначил телеведущую Тину Канделаки на пост исполняющей обязанности директора телеканала ТНТ, сообщается на сайте компании. Ранее о том, что Канделаки займет этот пост, рассказали источники РБК.
Ранее о том, что Канделаки займет этот пост, рассказали источники РБК.
С сентября 2021 года Канделаки курирует работу развлекательных телеканалов. Помимо ТНТ это ТВ-3, «Пятница», 2х2 и другие. Она также отвечает за развитие информагентства «Матч».
Накануне о перестановках на ТНТ сообщили источники РБК на медиарынке. Пост руководителя ТНТ покинул Роман Петренко. По словам собеседников РБК, из холдинга он при этом не уходит — ему предложили должность советника гендиректора «Газпром-Медиа» Александра Жарова.
Ранее источник РБК рассказывал, что Петренко был недоволен фактическим сокращением своих полномочий. Раньше он отвечал за финансовую политику ТНТ и косвенно за деятельность онлайн-кинотеатра Premier, но в последние месяцы его роль в холдинге существенно уменьшилась.
Туберкулезный некротизирующий токсин представляет собой НАД+ и НАДФ+ гликогидролазу с различными ферментативными свойствами
При инфицировании хозяина Mycobacterium tuberculosis выделяют туберкулезный некротизирующий токсин (TNT) в цитозоль инфицированных макрофагов, что приводит к гибели клетки-хозяина в результате некроптоза. TNT гидролизует NAD + в отсутствие какого-либо экзогенного кофактора, что классифицирует его как гликогидролазу β-NAD + .Однако у TNT отсутствует сходство последовательностей с другими ферментами, гидролизующими NAD + , и отсутствуют основные мотивы, участвующие в связывании и гидролизе NAD + этими ферментами. В этом исследовании мы использовали ЯМР для изучения ферментативной активности TNT и обнаружили, что TNT гидролизует NADP + так же быстро, как NAD + , но не расщепляет соответствующие восстановленные динуклеотиды. Эта активность ТНТ не ингибировалась АДФ-рибозой или никотинамидом, что указывает на низкое сродство ТНТ к этим продуктам реакции.Анализ селекции нетоксичных вариантов TNT в Escherichia coli выявил четыре из шести остатков в предсказанном кармане связывания NAD + и четыре остатка глицина, которые образуют колыбель непосредственно под сайтом связывания NAD + , законсервированная особенность в семейство белков TNT.
TNT гидролизует NAD + в отсутствие какого-либо экзогенного кофактора, что классифицирует его как гликогидролазу β-NAD + .Однако у TNT отсутствует сходство последовательностей с другими ферментами, гидролизующими NAD + , и отсутствуют основные мотивы, участвующие в связывании и гидролизе NAD + этими ферментами. В этом исследовании мы использовали ЯМР для изучения ферментативной активности TNT и обнаружили, что TNT гидролизует NADP + так же быстро, как NAD + , но не расщепляет соответствующие восстановленные динуклеотиды. Эта активность ТНТ не ингибировалась АДФ-рибозой или никотинамидом, что указывает на низкое сродство ТНТ к этим продуктам реакции.Анализ селекции нетоксичных вариантов TNT в Escherichia coli выявил четыре из шести остатков в предсказанном кармане связывания NAD + и четыре остатка глицина, которые образуют колыбель непосредственно под сайтом связывания NAD + , законсервированная особенность в семейство белков TNT. Сайт-направленный мутагенез остатков вблизи предполагаемого сайта связывания NAD + показал, что Phe 727 , Arg 757 и Arg 780 необходимы для гидролиза NAD + с помощью TNT.Эти результаты идентифицируют сайт связывания NAD + TNT. Наши результаты также показывают, что TNT представляет собой гликогидролазу NAD + со свойствами, отличными от свойств других бактериальных гликогидролаз. Поскольку многие из этих остатков сохраняются в семействе TNT, наши результаты позволяют лучше понять функцию более 300 гомологов TNT.
Сайт-направленный мутагенез остатков вблизи предполагаемого сайта связывания NAD + показал, что Phe 727 , Arg 757 и Arg 780 необходимы для гидролиза NAD + с помощью TNT.Эти результаты идентифицируют сайт связывания NAD + TNT. Наши результаты также показывают, что TNT представляет собой гликогидролазу NAD + со свойствами, отличными от свойств других бактериальных гликогидролаз. Поскольку многие из этих остатков сохраняются в семействе TNT, наши результаты позволяют лучше понять функцию более 300 гомологов TNT.
Ключевые слова: бактериальный токсин; кинетика ферментов; ферментативный механизм; взаимодействие хозяин-патоген; туберкулёз.
Новый сериал TNT «Уилл» вопиюще глуп, глэм-рок Шекспир
Вопрос «Что, если бы Уильям Шекспир действительно был бы действительно хорош собой?» уже получил окончательный ответ в оскароносном фильме 1998 года « Влюбленный Шекспир». Итак, Уилл , дебютировавший на канале TNT в понедельник вечером, пошел еще дальше. Что, если Шекспир на самом деле был очень красивым и тайным католическим шпионом и прототипом иконы глэм-рока и , страдающим посттравматическим стрессовым расстройством, вызванным наблюдением за тем, как его дядю публично выпотрошили? А что, если все это происходило в каком-то галлюциногенном стимпанковом лихорадочном сне База Лурмана о Лондоне 16-го века, где татуированные девицы из таверны носят дреды цвета фуксии, а гуляки врываются в исполнение «Идеального дня» Лу Рида?
Итак, Уилл , дебютировавший на канале TNT в понедельник вечером, пошел еще дальше. Что, если Шекспир на самом деле был очень красивым и тайным католическим шпионом и прототипом иконы глэм-рока и , страдающим посттравматическим стрессовым расстройством, вызванным наблюдением за тем, как его дядю публично выпотрошили? А что, если все это происходило в каком-то галлюциногенном стимпанковом лихорадочном сне База Лурмана о Лондоне 16-го века, где татуированные девицы из таверны носят дреды цвета фуксии, а гуляки врываются в исполнение «Идеального дня» Лу Рида?
Много чего происходит.И все же вся избыточная и неистовая энергия не может нести в себе сюжет, который полностью теряется во второстепенной сюжетной линии о Реформации и о том, как протестантский силовик Ричард Топклифф (которого здесь в ласковом стиле играет Юэн Бремнер из На игле ) охотился и пытали католиков. Уилл создан Крейгом Пирсом, соавтором Лурмана, который вместе с режиссером написал Ромео + Джульетта, Мулен Руж!, и Великий Гэтсби . Многие отличительные черты Лурмана — яркие, кричащие цвета; огромные театральные декорации; анахроничная музыка — придайте сериалу из 10 частей особое чувство стиля.Однако важно отметить, что Уилл , похоже, не слишком интересуется Шекспиром как драматургом. И возникающие в результате попытки впихнуть его в другие роли — знойного любовника, новоиспеченной знаменитости, упорного ренегата — часто приводят к сбивающим с толку впечатлениям от просмотра.
Многие отличительные черты Лурмана — яркие, кричащие цвета; огромные театральные декорации; анахроничная музыка — придайте сериалу из 10 частей особое чувство стиля.Однако важно отметить, что Уилл , похоже, не слишком интересуется Шекспиром как драматургом. И возникающие в результате попытки впихнуть его в другие роли — знойного любовника, новоиспеченной знаменитости, упорного ренегата — часто приводят к сбивающим с толку впечатлениям от просмотра.
В первом эпизоде Уилл (которого серьезно играет новичок Лори Дэвидсон) — мастер перчаток, который бросает свою жену и троих детей в Стратфорде-на-Эйвоне, чтобы искать счастья в качестве драматурга. Город, в который он прибывает, в этом наиболее заезженном из телевизионных тропов перегруженных деревенских мышей, — это Лондон через «Пираты Карибского моря »: зеленый экран и изобилующий пороком.Есть карманники, угри, публичные акты стоматологии, агрессивная неоновая цветовая палитра и даже заблудший верблюд. На мастера Шекспира, совершенно не в его лиге, охотится молодой уличный мальчишка (Лукас Рольфе), который крадет не только кошелек своей жертвы, но и осуждающее письмо, в котором Уилл изображается католиком.
Факты о жизни Шекспира настолько туманны, что Пирс вправе позволить себе вольности с его биографией. Но сюжет религиозного конфликта не просто утомителен — он также кажется явной попыткой внедрить ритуальное насилие и садизм в стиле Game of Thrones — в происходящее.Гораздо интереснее сцены, где Уилл почти случайно находит свое новое племя в театре Джеймса Бербеджа (Колм Мини). Среди них дети Бербеджа, Ричард (Маттиас Инвуд), прихорашивающийся актер, и Элис (Оливия ДеДжонг), которая помогает Уиллу структурировать и расшифровывать его пьесы и чье обаятельное обаяние помогает ему еще быстрее уйти от семьи, которую он оставил.
Есть также Кристофер Марлоу, необходимый в любом шекспировском биографическом фильме, интерпретируемый здесь Джейми Кэмпбеллом Бауэром как идол в стиле Марка Болана, который лечит свой писательский кризис оргиями, в которых его обхаживают флотилии обнаженных мужчин-моделей.Марлоу — сопровождающий Уилла в развратную изнанку елизаветинского Лондона и самые глупые сцены в сериале, а именно опиумную вакханалию, устроенную сэром Фрэнсисом Бэконом, которого представили как «нашего хозяина, отъявленного содомита». У Бауэра есть скулы и презрение, чтобы справиться с персонажем, но его вопиющие звериные аппетиты делают Уилла Дэвидсона по сравнению с ним довольно унылым, с широко открытыми глазами и приписывающими все свои лучшие реплики будущего из того, что люди говорят на вечеринках. Эти инста-цитаты о дивных новых мирах и пьесе появляются часто, но они не могут возвысить остальную часть сценария, наполненную безвкусицей.Католические трактаты, как говорят Уиллу, должны проникать в души своих читателей, как «пуля, выпущенная из смертоносного пистолета», но только тогда, когда они не оттаивают сердца людей, «как весна оттаивает могучую реку».
У Бауэра есть скулы и презрение, чтобы справиться с персонажем, но его вопиющие звериные аппетиты делают Уилла Дэвидсона по сравнению с ним довольно унылым, с широко открытыми глазами и приписывающими все свои лучшие реплики будущего из того, что люди говорят на вечеринках. Эти инста-цитаты о дивных новых мирах и пьесе появляются часто, но они не могут возвысить остальную часть сценария, наполненную безвкусицей.Католические трактаты, как говорят Уиллу, должны проникать в души своих читателей, как «пуля, выпущенная из смертоносного пистолета», но только тогда, когда они не оттаивают сердца людей, «как весна оттаивает могучую реку».
Странно то, что когда Уилл сосредотачивается на творческом процессе своего героя, это совсем неплохо. Импровизированная рэп-битва в первом эпизоде, исполненная пятистопным ямбом, похоже, вдохновлена Гамильтоном , но позволяет шоу задействовать самые интересные качества своего главного героя.Возможно, отказываясь уделять много внимания тому, как рождались шекспировские пьесы, шоу хотело избежать слишком похожего на Влюбленный Шекспир , хотя сцены с чернильно-черными ручками, царапающими пергамент, и лихорадочные свидания под сценой подразумевают обратное. Уилла можно простить за то, что он переосмыслил величайшего драматурга в истории как курящего наркотики суперзвезду глэм-рока. Но, к сожалению, не для того, чтобы отодвинуть его бесспорную гениальность на второстепенную сюжетную линию.
Уилла можно простить за то, что он переосмыслил величайшего драматурга в истории как курящего наркотики суперзвезду глэм-рока. Но, к сожалению, не для того, чтобы отодвинуть его бесспорную гениальность на второстепенную сюжетную линию.
Секреция и перенос токсина Mycobacterium tuberculosis
Мотивы ESX необходимы для стабильности CpnT и/или секреции TNT
M.туберкулез CpnT не содержит канонического сигнального пептида Sec, в отличие от любых других известных белков внешней мембраны (дополнительный рисунок 1). Поскольку CpnT содержит три мотива YXXXE, которые, как ранее было показано, необходимы для секреции определенных субстратов ESX-1 18 , мы стремились исследовать, необходимы ли они для экспорта CpnT с помощью Mtb. Эти мотивы расположены в положениях Y88, Y356 и Y741 и далее обозначаются как мотивы Y1, Y2 и Y3 соответственно (рис.1а, дополнительный рис. 1). С этой целью мы использовали штамм Mtb ML2016 (дополнительная таблица 1), в котором отсутствует весь оперон cpnT , и дополнили этот штамм с помощью векторов интегративной экспрессии, несущих оперон cpnT , либо с геном wt cpnT , либо с геном . cpnT , кодирующие белки, в которых консервативные тирозины каждого Y-мотива мутированы в аланин (далее Y1*, Y2* и Y3*). Хотя экспрессия оперона, продуцирующего CpnT, CpnT Y2* и Y3* дикого типа, приводила к сходным уровням белка, для CpnT Y1* белок не был обнаружен (рис.1б). Сходные уровни фактора иммунитета к TNT (IFT) у всех штаммов продемонстрировали, что все гены оперонов транслировались одинаково (рис. 1b), и, следовательно, указывали на то, что отсутствие белка CpnT-Y1* является результатом посттрансляционного события. Флуоресцентная микроскопия штаммов Mtb, несущих различные аллели cpnT , с использованием антител против TNT показала, что домен TNT доступен на клеточной поверхности Mtb (рис. 1c, дополнительная рис. 2), как показано ранее 6, 8 . Как и ожидалось, домен TNT не был обнаружен в Mtb, лишенном оперона cpnT или кодирующего CpnT с мутированным мотивом ESX-Y1*, но, что удивительно, TNT также не был обнаружен на клеточной поверхности в Mtb-кодирующем CpnT с мутированным Мотив ESX-Y3* (рис.
cpnT , кодирующие белки, в которых консервативные тирозины каждого Y-мотива мутированы в аланин (далее Y1*, Y2* и Y3*). Хотя экспрессия оперона, продуцирующего CpnT, CpnT Y2* и Y3* дикого типа, приводила к сходным уровням белка, для CpnT Y1* белок не был обнаружен (рис.1б). Сходные уровни фактора иммунитета к TNT (IFT) у всех штаммов продемонстрировали, что все гены оперонов транслировались одинаково (рис. 1b), и, следовательно, указывали на то, что отсутствие белка CpnT-Y1* является результатом посттрансляционного события. Флуоресцентная микроскопия штаммов Mtb, несущих различные аллели cpnT , с использованием антител против TNT показала, что домен TNT доступен на клеточной поверхности Mtb (рис. 1c, дополнительная рис. 2), как показано ранее 6, 8 . Как и ожидалось, домен TNT не был обнаружен в Mtb, лишенном оперона cpnT или кодирующего CpnT с мутированным мотивом ESX-Y1*, но, что удивительно, TNT также не был обнаружен на клеточной поверхности в Mtb-кодирующем CpnT с мутированным Мотив ESX-Y3* (рис. 1c, дополнительный рисунок 2), хотя уровни белка и мембранная ассоциация в экспериментах по субклеточному фракционированию CpnT Y3* аналогичны таковым у CpnT дикого типа (рис. 1b, дополнительный рисунок 3). Чтобы определить относительное количество TNT на поверхности клеток, мы измерили флуоресценцию отдельных клеток Mtb с помощью проточной цитометрии, как показано ранее 5, 8 . Результаты этого количественного анализа согласуются с экспериментами по флуоресцентной микроскопии: штаммы Mtb с мутированными мотивами ESX Y1* и Y3* не имели обнаруживаемого на поверхности TNT, в то время как штамм Mtb с мутированным мотивом ESX Y2* имеет уровни дикого типа TNT, экспонированные на клеточная поверхность Mtb (рис.1г).
1c, дополнительный рисунок 2), хотя уровни белка и мембранная ассоциация в экспериментах по субклеточному фракционированию CpnT Y3* аналогичны таковым у CpnT дикого типа (рис. 1b, дополнительный рисунок 3). Чтобы определить относительное количество TNT на поверхности клеток, мы измерили флуоресценцию отдельных клеток Mtb с помощью проточной цитометрии, как показано ранее 5, 8 . Результаты этого количественного анализа согласуются с экспериментами по флуоресцентной микроскопии: штаммы Mtb с мутированными мотивами ESX Y1* и Y3* не имели обнаруживаемого на поверхности TNT, в то время как штамм Mtb с мутированным мотивом ESX Y2* имеет уровни дикого типа TNT, экспонированные на клеточная поверхность Mtb (рис.1г).
Уровни белка CpnT очень низки в Mtb, выращенном in vitro, но сильно повышены в Mtb после инфицирования макрофагов 6 . Следовательно, мы использовали клеточную линию макрофагов человека THP-1, чтобы проверить, влияют ли мотивы YXXXE на секрецию CpnT/TNT таким же образом, как и in vitro. Большие количества TNT в цитозоле инфицированных макрофагов THP-1 наблюдались при экспрессии оперона wt cpnT или оперона с cpnT-Y2* , в то время как для cpnT-Y1* и секретируемый TNT не обнаруживался. cpnT-Y3* (рис.1e, f, дополнительный рисунок 4), что соответствует результатам in vitro. В совокупности эти эксперименты установили, что мутация мотивов ESX Y1 и Y3 предотвращает достижение CpnT клеточной поверхности Mtb. Отсутствие белка CpnT с мутантным мотивом ESX Y1, вероятно, связано с деградацией белка, вероятно, вызванной накоплением и агрегацией белка, когда транслокация белка нарушена 19 . Напротив, уровни белка и мембранная ассоциация CpnT ESX Y3* сходны с таковыми у CpnT дикого типа, что указывает на существенную роль мотива ESX Y3 в транслокации через внешнюю мембрану.Однако для обоих мутантов мы не можем исключить вредное влияние точечных мутаций Y88A и Y741A на стабильность белка CpnT.
Большие количества TNT в цитозоле инфицированных макрофагов THP-1 наблюдались при экспрессии оперона wt cpnT или оперона с cpnT-Y2* , в то время как для cpnT-Y1* и секретируемый TNT не обнаруживался. cpnT-Y3* (рис.1e, f, дополнительный рисунок 4), что соответствует результатам in vitro. В совокупности эти эксперименты установили, что мутация мотивов ESX Y1 и Y3 предотвращает достижение CpnT клеточной поверхности Mtb. Отсутствие белка CpnT с мутантным мотивом ESX Y1, вероятно, связано с деградацией белка, вероятно, вызванной накоплением и агрегацией белка, когда транслокация белка нарушена 19 . Напротив, уровни белка и мембранная ассоциация CpnT ESX Y3* сходны с таковыми у CpnT дикого типа, что указывает на существенную роль мотива ESX Y3 в транслокации через внешнюю мембрану.Однако для обоих мутантов мы не можем исключить вредное влияние точечных мутаций Y88A и Y741A на стабильность белка CpnT.
Система ESX-4 необходима для доступа к поверхности CpnT в
M. tuberculosis
tuberculosis Эксперименты по доступности поверхности с использованием CpnT с мутациями в предполагаемых мотивах ESX согласуются с ролью системы секреции типа VII в экспорте CpnT, но не являются окончательными. Поэтому мы исследовали роль систем ESX в экспорте CpnT и секреции TNT.Хотя мотивы YXXXE необходимы для секреции субстратов ESX-1 и ESX-5 18 , предыдущие эксперименты показали, что экспорт CpnT и поверхностная локализация TNT в Mtb не зависят от системы секреции ESX-1 6 . Эти результаты были получены с использованием авирулентного мутанта Mtb mc 2 6206, что указывает на то, что экспорт CpnT и секреция TNT существенно не изменяются при делеции генов leuCD и panCD (дополнительная таблица 1).Это было экспериментально подтверждено в экспериментах по заражению макрофагов с использованием вирулентного штамма Mtb h47Rv, который показал аналогичную секрецию TNT (дополнительная рис. 5) и цитотоксичность (дополнительная рис. 6) в инфицированных макрофагах по сравнению с Mtb mc 2 6206. Эти результаты согласуются с предыдущими исследованиями, демонстрирующими, что механизм индуцированной TNT гибели клеток одинаков у вирулентных и авирулентных штаммов Mtb 7, 20 , а также с недавним исследованием, демонстрирующим, что вирулентный штамм Mtb h47Rv и авирулентный Mtb Δ leuD Δ panCD имеют одинаковую скорость репликации in vitro и в макрофагах и вызывают сходный цитокиновый ответ 21 .Следовательно, мы использовали Mtb mc 2 6206 для получения мутантов с нефункциональными системами ESX-2 и ESX-4 путем создания немаркированных внутрирамочных делеций генов eccC2 и eccC4 соответственно (дополнительная рис. 7a). – г). Гены eccC кодируют специфические АТФазы, которые необходимы для функционирования их соответствующей системы ESX 22, 23 . Таким образом, делеции в генах eccC2 и eccC4 привели к появлению штаммов Mtb ML2691 и ML2690 (дополнительная таблица 1) с нефункциональными системами секреции ESX-2 и ESX-4 соответственно, которые отныне обозначаются как Δ .
6) в инфицированных макрофагах по сравнению с Mtb mc 2 6206. Эти результаты согласуются с предыдущими исследованиями, демонстрирующими, что механизм индуцированной TNT гибели клеток одинаков у вирулентных и авирулентных штаммов Mtb 7, 20 , а также с недавним исследованием, демонстрирующим, что вирулентный штамм Mtb h47Rv и авирулентный Mtb Δ leuD Δ panCD имеют одинаковую скорость репликации in vitro и в макрофагах и вызывают сходный цитокиновый ответ 21 .Следовательно, мы использовали Mtb mc 2 6206 для получения мутантов с нефункциональными системами ESX-2 и ESX-4 путем создания немаркированных внутрирамочных делеций генов eccC2 и eccC4 соответственно (дополнительная рис. 7a). – г). Гены eccC кодируют специфические АТФазы, которые необходимы для функционирования их соответствующей системы ESX 22, 23 . Таким образом, делеции в генах eccC2 и eccC4 привели к появлению штаммов Mtb ML2691 и ML2690 (дополнительная таблица 1) с нефункциональными системами секреции ESX-2 и ESX-4 соответственно, которые отныне обозначаются как Δ .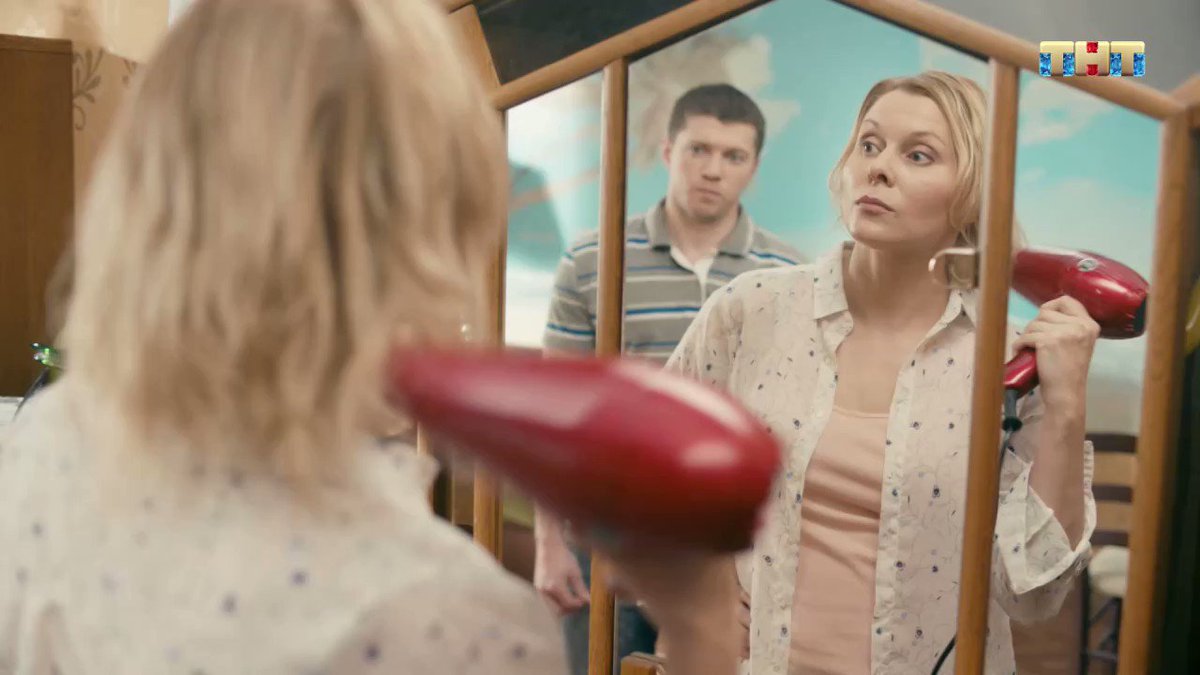 штаммы esx-2 и Δ esx-4 .Поскольку ген eccC2 находится в непосредственной близости от локуса esx-1 , мы исследовали секрецию субстрата ESX-1 CFP-10 и показали, что уровни белка CFP-10 идентичны в фильтратах культур . esx-2- и штаммы с дефицитом esx-4- и его родительский штамм Mtb mc 2 6206, но CFP-10 не был обнаружен в мутанте Δ esx-1 (дополнительный рисунок 8). Этот эксперимент показал, что делеция eccC2 не изменила активность системы ESX-1.В последующих экспериментах в качестве отрицательного контроля использовали делеционный мутант esx-1 Mtb mc 2 6230 (дополнительная таблица 1). Флуоресцентная микроскопия показала, что TNT-домен CpnT экспонирован на клеточной поверхности во всех штаммах Mtb, за исключением штамма, лишенного функциональной системы ESX-4, и, как и ожидалось, в мутанте с делецией оперона cpnT (рис. 2a, b, Дополнительный рис. 9). Доступность поверхности TNT была восстановлена, когда локус esx-4 на космиде I60 (дополнительная таблица 2) был введен в мутант Mtb Δ esx-4 (рис.
штаммы esx-2 и Δ esx-4 .Поскольку ген eccC2 находится в непосредственной близости от локуса esx-1 , мы исследовали секрецию субстрата ESX-1 CFP-10 и показали, что уровни белка CFP-10 идентичны в фильтратах культур . esx-2- и штаммы с дефицитом esx-4- и его родительский штамм Mtb mc 2 6206, но CFP-10 не был обнаружен в мутанте Δ esx-1 (дополнительный рисунок 8). Этот эксперимент показал, что делеция eccC2 не изменила активность системы ESX-1.В последующих экспериментах в качестве отрицательного контроля использовали делеционный мутант esx-1 Mtb mc 2 6230 (дополнительная таблица 1). Флуоресцентная микроскопия показала, что TNT-домен CpnT экспонирован на клеточной поверхности во всех штаммах Mtb, за исключением штамма, лишенного функциональной системы ESX-4, и, как и ожидалось, в мутанте с делецией оперона cpnT (рис. 2a, b, Дополнительный рис. 9). Доступность поверхности TNT была восстановлена, когда локус esx-4 на космиде I60 (дополнительная таблица 2) был введен в мутант Mtb Δ esx-4 (рис. 2а, б, дополнительный рис. 9). Важно отметить, что количественный анализ интактных клеток Mtb с помощью проточной цитометрии с использованием TNT-антитела, как описано выше, подтвердил полное отсутствие доступного на поверхности CpnT у мутанта Mtb с нефункциональной системой ESX4. Экспрессия локуса esx-4 на космиде I60 полностью восстановила доступность поверхности CpnT в Mtb (рис. 2c). Эксперименты по субклеточному фракционированию показали большую часть белка CpnT в мембранной фракции штамма Mtb с дефицитом esx-4 (дополнительная рис.10). Поскольку у esx-4 -дефицитных Mtb не наблюдалось воздействия на поверхность TNT-домена, такое совместное фракционирование могло быть результатом либо нефункциональной интеграции CpnT в мембрану, либо агрегатов неправильно свернутых CpnT из-за нарушения экспорта, как это наблюдалось. для других белков 19 . В совокупности эти эксперименты демонстрируют, что система секреции ESX-4 необходима для транслокации CpnT на клеточную поверхность Mtb.
2а, б, дополнительный рис. 9). Важно отметить, что количественный анализ интактных клеток Mtb с помощью проточной цитометрии с использованием TNT-антитела, как описано выше, подтвердил полное отсутствие доступного на поверхности CpnT у мутанта Mtb с нефункциональной системой ESX4. Экспрессия локуса esx-4 на космиде I60 полностью восстановила доступность поверхности CpnT в Mtb (рис. 2c). Эксперименты по субклеточному фракционированию показали большую часть белка CpnT в мембранной фракции штамма Mtb с дефицитом esx-4 (дополнительная рис.10). Поскольку у esx-4 -дефицитных Mtb не наблюдалось воздействия на поверхность TNT-домена, такое совместное фракционирование могло быть результатом либо нефункциональной интеграции CpnT в мембрану, либо агрегатов неправильно свернутых CpnT из-за нарушения экспорта, как это наблюдалось. для других белков 19 . В совокупности эти эксперименты демонстрируют, что система секреции ESX-4 необходима для транслокации CpnT на клеточную поверхность Mtb. Этот результат также установил, что CpnT является первым белком наружной мембраны, зависящим от системы секреции типа VII у Mtb.
Этот результат также установил, что CpnT является первым белком наружной мембраны, зависящим от системы секреции типа VII у Mtb.
a Обнаружение поверхностно-доступного CpnT в Mtb с помощью флуоресцентной микроскопии. Штаммы Mtb метили DMN-трегалозой (зеленый) и окрашивали поликлональными анти-ТНТ и вторичными антителами Alexa Fluor-594 (красный). b Количественное определение TNT-положительных клеток Mtb по изображениям, показанным в a . Клетки Mtb оценивали как TNT-положительные, когда наблюдали красный сигнал по сравнению с Mtb Δ cpnT op . c Доступность поверхности CpnT в Mtb с помощью проточной цитометрии. Штаммы Mtb окрашивали поликлональными анти-ТНТ и вторичными антителами Alexa Fluor-488. Средняя флуоресценция клеток Mtb отображается на гистограммах. d Поверхностная доступность CpnT делеционного мутанта Mtb h47Rv esx-3 и контрольных штаммов с помощью флуоресцентной микроскопии. Процедуру окрашивания проводили как в и . e Количественное определение TNT-положительных клеток Mtb по изображениям, показанным в d .Клетки Mtb оценивали как TNT-положительные, когда наблюдали красный сигнал по сравнению с Mtb Δ cpnT . f Выход из строя функциональной системы ESX-5 в Mtb Erdman. Мутантные штаммы wt и eccD 5 Tet-OFF выращивали в присутствии/отсутствии гидрохлорида ангидротетрациклина (ATc; 100 нг/мл). Белки из цельноклеточных лизатов и культуральных фильтратов анализировали с помощью иммуноблоттинга для обнаружения EccD 5 , PPE41 и CpnT со специфическими антителами. г Поверхностная доступность CpnT в Mtb Erdman, лишенном функциональной системы Esx-5, с помощью флуоресцентной микроскопии. Процедуру окрашивания проводили как в и . Штаммы wt и Δ cpnT на фоне h47Rv использовали в качестве положительного и отрицательного контроля соответственно. h TNT-положительные клетки Mtb из изображений, показанных в g , количественно определяли, как описано в e .
Процедуру окрашивания проводили как в и . e Количественное определение TNT-положительных клеток Mtb по изображениям, показанным в d .Клетки Mtb оценивали как TNT-положительные, когда наблюдали красный сигнал по сравнению с Mtb Δ cpnT . f Выход из строя функциональной системы ESX-5 в Mtb Erdman. Мутантные штаммы wt и eccD 5 Tet-OFF выращивали в присутствии/отсутствии гидрохлорида ангидротетрациклина (ATc; 100 нг/мл). Белки из цельноклеточных лизатов и культуральных фильтратов анализировали с помощью иммуноблоттинга для обнаружения EccD 5 , PPE41 и CpnT со специфическими антителами. г Поверхностная доступность CpnT в Mtb Erdman, лишенном функциональной системы Esx-5, с помощью флуоресцентной микроскопии. Процедуру окрашивания проводили как в и . Штаммы wt и Δ cpnT на фоне h47Rv использовали в качестве положительного и отрицательного контроля соответственно. h TNT-положительные клетки Mtb из изображений, показанных в g , количественно определяли, как описано в e . Штамм Δ esx-1 имеет делецию области RD1 и лишен генов panCD .Данные представлены как среднее ± SEM по крайней мере двух независимых экспериментов ( n ≥ 2), и показаны репрезентативные изображения/пятна. Звездочками отмечены значимые различия (* p значение ≤ 0,05, ** p значение ≤ 0,01, *** значение ≤ 0,001, **** p значение ≤ 0,0001, рассчитанные с использованием однофакторного дисперсионного анализа с поправкой Даннета ) по сравнению с соответствующим штаммом дикого типа. Исходные данные предоставляются в файле исходных данных.
Штамм Δ esx-1 имеет делецию области RD1 и лишен генов panCD .Данные представлены как среднее ± SEM по крайней мере двух независимых экспериментов ( n ≥ 2), и показаны репрезентативные изображения/пятна. Звездочками отмечены значимые различия (* p значение ≤ 0,05, ** p значение ≤ 0,01, *** значение ≤ 0,001, **** p значение ≤ 0,0001, рассчитанные с использованием однофакторного дисперсионного анализа с поправкой Даннета ) по сравнению с соответствующим штаммом дикого типа. Исходные данные предоставляются в файле исходных данных.
Системы ESX-3 и ESX-5 не участвуют в экспорте CpnT в
M.туберкулез Чтобы изучить роль системы ESX-3 в экспорте CpnT, мы использовали делеционный мутант esx-3 Mtb mc 2 7788 13 , производное Mtb h47Rv (дополнительная таблица 1). Флуоресцентная микроскопия показала, что экспозиция домена TNT на клеточной поверхности у Mtb h47Rv и штамма Δ esx-3 идентична, в отличие от таковой у делеционного мутанта cpnT (рис. 2d, e, дополнительная рис. 11), демонстрируя, что система ESX-3 не участвует в экспорте CpnT в Mtb.
2d, e, дополнительная рис. 11), демонстрируя, что система ESX-3 не участвует в экспорте CpnT в Mtb.
Чтобы определить, зависит ли экспорт CpnT от системы ESX-5, мы использовали условный мутант Mtb Erdman eccD 5 TET-OFF, который истощает трансмембранный канал EccD 5 в присутствии ангидротетрациклина (ATc) 24 . Использование этого штамма оправдано тем, что оперон cpnT , локус esx-4 и все консервативные гены локуса esx-5 идентичны у Mtb h47Rv и Mtb Erdman.Мы наблюдали 14-кратное снижение уровней EccD 5 у условного мутанта eccD 5 , выращенного в присутствии ATc (рис. 2f), что согласуется с предыдущими результатами 24 . Секреция субстрата ESX-5 PPE41 в культуральный фильтрат была полностью блокирована в присутствии ATc (рис. 2f), что свидетельствует о том, что система ESX-5 не функционирует у кондиционированного eccD 5 мутанта в условиях репрессии. условий и что низкие обнаруживаемые уровни EccD 5 недостаточны для поддержки функции системы ESX-5.Уровни CpnT в присутствии ATc у условного мутанта eccD 5 остались неизменными (рис. 2f). Чтобы выяснить, играет ли система ESX-5 роль в функциональной интеграции CpnT во внешней мембране, мы оценили экспозицию TNT на поверхности клеток Mtb с помощью флуоресцентной микроскопии. Как и ожидалось, TNT обнаруживался на поверхности Mtb h47Rv, но не у мутанта Δ cpnT , который использовался в качестве положительного и отрицательного контроля соответственно (рис.2g, h, дополнительный рис. 12). Важно отметить, что добавление ATc не ухудшило доступность CpnT на поверхности у мутанта eccD 5 -Tet-OFF (рис. 2g, h, дополнительная рис. 12), демонстрируя, что система секреции ESX-5 не участвует в экспорте, интеграции с наружной мембраной и экспонировании CpnT на поверхности Mtb. Взятые вместе с нашими предыдущими результатами, эти эксперименты демонстрируют, что ESX-4 является единственной системой секреции типа VII, необходимой для экспорта CpnT и экспонирования на поверхности его домена TNT в Mtb.
условий и что низкие обнаруживаемые уровни EccD 5 недостаточны для поддержки функции системы ESX-5.Уровни CpnT в присутствии ATc у условного мутанта eccD 5 остались неизменными (рис. 2f). Чтобы выяснить, играет ли система ESX-5 роль в функциональной интеграции CpnT во внешней мембране, мы оценили экспозицию TNT на поверхности клеток Mtb с помощью флуоресцентной микроскопии. Как и ожидалось, TNT обнаруживался на поверхности Mtb h47Rv, но не у мутанта Δ cpnT , который использовался в качестве положительного и отрицательного контроля соответственно (рис.2g, h, дополнительный рис. 12). Важно отметить, что добавление ATc не ухудшило доступность CpnT на поверхности у мутанта eccD 5 -Tet-OFF (рис. 2g, h, дополнительная рис. 12), демонстрируя, что система секреции ESX-5 не участвует в экспорте, интеграции с наружной мембраной и экспонировании CpnT на поверхности Mtb. Взятые вместе с нашими предыдущими результатами, эти эксперименты демонстрируют, что ESX-4 является единственной системой секреции типа VII, необходимой для экспорта CpnT и экспонирования на поверхности его домена TNT в Mtb.
Система ESX-4 участвует в транспорте EsxF на клеточную поверхность
M.tuberculosis Небольшие белки WXG100 EsxE и EsxF кодируются в опероне cpnT (рис. 1а) и образуют трансмембранный комплекс пор во внешней мембране, который необходим для экспорта CpnT и доступности поверхности TNT для Mtb 8 . Учитывая требование системы ESX-4 для экспорта CpnT в Mtb (рис. 2), мы задались вопросом, опосредован ли также экспорт комплекса EsxEF системой ESX-4.Чтобы проверить эту гипотезу, мы исследовали воздействие EsxF на поверхность Mtb с помощью флуоресцентной микроскопии и EsxF-специфических антител 8 . EsxF четко обнаруживается на клеточной поверхности в родительском штамме Mtb mc 2 6206, но не в штамме Mtb, в котором отсутствует оперон cpnT (рис. 3a, b, дополнительная рис. 13), что согласуется с предыдущими результатами 8 . Количественный анализ показал, что количество экспонированного на поверхности EsxF снижено примерно на 50% у штамма Mtb с нефункциональной системой ESX-4 по сравнению с исходным штаммом (рис. 3b), демонстрируя важную роль системы ESX-4 в экспорте и экспозиции комплекса EsxE-EsxF на поверхности. Остаточный поверхностно-доступный EsxF в мутанте Mtb с делецией esx-4 указывает на то, что в дополнение к ESX-4 существуют альтернативные пути для экспорта EsxF и поверхностной транслокации.
3b), демонстрируя важную роль системы ESX-4 в экспорте и экспозиции комплекса EsxE-EsxF на поверхности. Остаточный поверхностно-доступный EsxF в мутанте Mtb с делецией esx-4 указывает на то, что в дополнение к ESX-4 существуют альтернативные пути для экспорта EsxF и поверхностной транслокации.
a Обнаружение поверхностно-доступного EsxF указанных штаммов Mtb с помощью флуоресцентной микроскопии.Штаммы Mtb метили метаболическим красителем DMN-трегалозой (зеленый) и окрашивали поликлональными антителами против EsxF и вторичными антителами Alexa Fluor-594 (красный). Желтый цвет указывает на совместную локализацию EsxF с бактериальной клеткой. b Количественное определение EsxF-положительных клеток Mtb по изображениям, показанным в a . Клетки Mtb оценивали как EsxF-положительные, когда наблюдали красный сигнал по сравнению с мутантным делеционным опероном Mtb cpnT (Δ cpnT op ). Данные представлены как среднее ± SEM трех независимых экспериментов ( n = 3), и показаны репрезентативные изображения. Звездочками отмечены значимые различия (* p значение ≤ 0,05, ** p значение ≤ 0,01, *** p значение ≤ 0,001, **** p значение ≤ 0,0001, рассчитанные с использованием одностороннего ANOVA с поправкой Даннета) по сравнению со штаммом Mtb mc 2 6206. Исходные данные предоставляются в файле исходных данных.
Данные представлены как среднее ± SEM трех независимых экспериментов ( n = 3), и показаны репрезентативные изображения. Звездочками отмечены значимые различия (* p значение ≤ 0,05, ** p значение ≤ 0,01, *** p значение ≤ 0,001, **** p значение ≤ 0,0001, рассчитанные с использованием одностороннего ANOVA с поправкой Даннета) по сравнению со штаммом Mtb mc 2 6206. Исходные данные предоставляются в файле исходных данных.
Доставка CpnT в цитозоль макрофагов, инфицированных
M.tuberculosis зависит от систем ESX-1, ESX-2 и ESX-4 Так как система ESX-1 необходима для пермеабилизации фагосом 9, 25 и транслокации ТНТ в цитозоль макрофагов, инфицированных Mtb 6 , мы задались вопросом, могут ли другие системы ESX также участвовать в этом процессе. Для проверки этой гипотезы мы определили способность мутантных штаммов Mtb esx секретировать ТНТ в цитозоль инфицированных макрофагов. С этой целью мы визуализировали TNT в макрофагах THP-1, инфицированных различными штаммами Mtb, с помощью флуоресцентной микроскопии после селективной пермеабилизации детергентами дигитонином (только плазматическая мембрана) и Triton X-100 (плазма и фагосомальные мембраны), как описано ранее 6 , 26 .В качестве контроля использовали антитело, специфичное к комплексу антигена 85 (Ag85), которое обнаруживается в значительных количествах в супернатантах культур Mtb 27 . В экспериментах по заражению макрофагов Ag85 был обнаружен только тогда, когда макрофаги были пермеабилизированы Тритоном X-100, но не дигитонином (рис. 4a, дополнительная рис. 14), демонстрируя, что Ag85 содержится в фагосоме, как мы ранее описали 6 . Напротив, TNT секретируется Mtb mc 2 6206 в цитозоль инфицированных макрофагов THP-1 в соответствии с предыдущими публикациями 6, 7 .Как и ожидалось, мутант esx-1 продуцировал ТНТ, но он был ограничен фагосомой и не достигал цитозоля инфицированных макрофагов (рис.
С этой целью мы визуализировали TNT в макрофагах THP-1, инфицированных различными штаммами Mtb, с помощью флуоресцентной микроскопии после селективной пермеабилизации детергентами дигитонином (только плазматическая мембрана) и Triton X-100 (плазма и фагосомальные мембраны), как описано ранее 6 , 26 .В качестве контроля использовали антитело, специфичное к комплексу антигена 85 (Ag85), которое обнаруживается в значительных количествах в супернатантах культур Mtb 27 . В экспериментах по заражению макрофагов Ag85 был обнаружен только тогда, когда макрофаги были пермеабилизированы Тритоном X-100, но не дигитонином (рис. 4a, дополнительная рис. 14), демонстрируя, что Ag85 содержится в фагосоме, как мы ранее описали 6 . Напротив, TNT секретируется Mtb mc 2 6206 в цитозоль инфицированных макрофагов THP-1 в соответствии с предыдущими публикациями 6, 7 .Как и ожидалось, мутант esx-1 продуцировал ТНТ, но он был ограничен фагосомой и не достигал цитозоля инфицированных макрофагов (рис. 4a–c, дополнительная рис. 14). Этот фенотип наблюдался ранее 6 и согласуется с известной ролью ESX-1 в пермеабилизации фагосомальной мембраны 10, 28, 29 . ТНТ не обнаруживался в цитозоле или фагосомах макрофагов, инфицированных штаммом с дефицитом esx-4 (рис. 4a–c, дополнительная рис.14). Дополнение космидой I60 (дополнительная таблица 2), кодирующей функциональную систему ESX-4, восстанавливало фенотип дикого типа, подтверждая описанные выше эксперименты in vitro, показывающие, что система ESX-4 необходима для экспорта CpnT и воздействия на поверхность (рис. 2). Неожиданно мы не обнаружили ТНТ в цитозоле макрофагов, инфицированных мутантом esx-2 после пермеабилизации дигитонином. Однако пермеабилизация с помощью Triton X-100 выявила ТНТ вокруг клеток с дефицитом Mtb esx-2 (рис.4a–c, дополнительный рисунок 14). Этот результат предполагает, что TNT секретировался, но был захвачен фагосомой Mtb esx-2 -дефицитного штамма, подобно штамму Δ esx-1 .
4a–c, дополнительная рис. 14). Этот фенотип наблюдался ранее 6 и согласуется с известной ролью ESX-1 в пермеабилизации фагосомальной мембраны 10, 28, 29 . ТНТ не обнаруживался в цитозоле или фагосомах макрофагов, инфицированных штаммом с дефицитом esx-4 (рис. 4a–c, дополнительная рис.14). Дополнение космидой I60 (дополнительная таблица 2), кодирующей функциональную систему ESX-4, восстанавливало фенотип дикого типа, подтверждая описанные выше эксперименты in vitro, показывающие, что система ESX-4 необходима для экспорта CpnT и воздействия на поверхность (рис. 2). Неожиданно мы не обнаружили ТНТ в цитозоле макрофагов, инфицированных мутантом esx-2 после пермеабилизации дигитонином. Однако пермеабилизация с помощью Triton X-100 выявила ТНТ вокруг клеток с дефицитом Mtb esx-2 (рис.4a–c, дополнительный рисунок 14). Этот результат предполагает, что TNT секретировался, но был захвачен фагосомой Mtb esx-2 -дефицитного штамма, подобно штамму Δ esx-1 . Дополнение esx-2 -дефицитного штамма функциональной системой ESX-2 с использованием космиды I106 (дополнительная таблица 2) вернуло этот фенотип и обеспечило доступ TNT к цитозолю (рис. 4a–c, дополнительная рис. 14). В связи с этим важно отметить, что штаммы Mtb с дефицитом esx-2 и esx-4- сохранили свою способность продуцировать фтиоцеролдимикоцеросат (PDIM) (дополнительная фигура 7e), гликолипид клеточной стенки Mtb, известный как часто теряется во время культивирования in vitro и выполняет важные функции в патогенезе Mtb, такие как повреждение и разрыв фагосомной мембраны, защита от зависимого от оксида азота уничтожения 30, 31 и функция белка наружной мембраны 32 .Штамм Mtb h47Rv Δ esx-3 и кондициональный мутант Mtb esx-5 в условиях репрессии, нарушающих функцию системы ESX-5 (рис. 2е–з), секретировали ТНТ в цитозоль аналогично как исходный штамм (рис. 4e, f, дополнительные рисунки 15, 16). Эти результаты согласуются с экспериментами in vitro и исключают вклад систем ESX-3 и ESX-5 в секрецию TNT и перенос Mtb.
Дополнение esx-2 -дефицитного штамма функциональной системой ESX-2 с использованием космиды I106 (дополнительная таблица 2) вернуло этот фенотип и обеспечило доступ TNT к цитозолю (рис. 4a–c, дополнительная рис. 14). В связи с этим важно отметить, что штаммы Mtb с дефицитом esx-2 и esx-4- сохранили свою способность продуцировать фтиоцеролдимикоцеросат (PDIM) (дополнительная фигура 7e), гликолипид клеточной стенки Mtb, известный как часто теряется во время культивирования in vitro и выполняет важные функции в патогенезе Mtb, такие как повреждение и разрыв фагосомной мембраны, защита от зависимого от оксида азота уничтожения 30, 31 и функция белка наружной мембраны 32 .Штамм Mtb h47Rv Δ esx-3 и кондициональный мутант Mtb esx-5 в условиях репрессии, нарушающих функцию системы ESX-5 (рис. 2е–з), секретировали ТНТ в цитозоль аналогично как исходный штамм (рис. 4e, f, дополнительные рисунки 15, 16). Эти результаты согласуются с экспериментами in vitro и исключают вклад систем ESX-3 и ESX-5 в секрецию TNT и перенос Mtb. В совокупности эти результаты показывают, что системы ESX-1 и ESX-2 не могут заменять друг друга и что активность обеих систем ESX необходима для переноса TNT в цитозоль макрофагов, инфицированных Mtb.
В совокупности эти результаты показывают, что системы ESX-1 и ESX-2 не могут заменять друг друга и что активность обеих систем ESX необходима для переноса TNT в цитозоль макрофагов, инфицированных Mtb.
a Секреция ТНТ в цитозоль Mtb-инфицированных макрофагов. Указанные штаммы Mtb были помечены метаболическим красителем DMN-трегалоза (зеленый) и использованы для заражения макрофагов THP-1 при множественности заражения 10:1. Через 48 ч после заражения макрофаги пермеабилизировали дигитонином, чтобы обеспечить доступ антител к цитоплазме, или тритоном Х-100, чтобы получить доступ к внутриклеточным компартментам.Затем клетки окрашивали антителами против TNT или Ag85 и вторичными антителами Alexa Fluor-594 (красные). Ядра макрофагов окрашивали DAPI. b , c Количественное определение TNT-положительных макрофагов после пермеабилизации дигитонином ( b ) или Triton X-100 ( c ) по изображениям, показанным в a . Макрофаги считали ТНТ-позитивными, когда наблюдали отчетливые красные точки по сравнению с мутантным делеционным опероном Mtb cpnT (Δ cpnT op ). d Жизнеспособность клеток Mtb-инфицированных макрофагов измеряли по общему содержанию АТФ с помощью набора для определения люминесцентного АТФ. e Секреция ТНТ в цитозоль инфицированных макрофагов в штаммах Mtb h47Rv. Заражение макрофагами и процедуру окрашивания проводили, как описано в и . f Секреция ТНТ в цитозоль инфицированных макрофагов штаммов Mtb h47Rv и Erdman. Заражение макрофагами и процедуру окрашивания проводили, как описано в и .По показаниям бактерии выращивали в 7H9 + 100 нг/мл ангидротетрациклина (ATc) для предварительного истощения EccD5 перед инфицированием, затем ATc сохраняли в культуральной среде во время инфицирования. Штамм Δ esx-1 имеет делецию области RD1 и лишен генов panCD . Данные представлены как среднее ± SEM по крайней мере двух независимых экспериментов ( n ≥ 2), и показаны репрезентативные изображения.
Макрофаги считали ТНТ-позитивными, когда наблюдали отчетливые красные точки по сравнению с мутантным делеционным опероном Mtb cpnT (Δ cpnT op ). d Жизнеспособность клеток Mtb-инфицированных макрофагов измеряли по общему содержанию АТФ с помощью набора для определения люминесцентного АТФ. e Секреция ТНТ в цитозоль инфицированных макрофагов в штаммах Mtb h47Rv. Заражение макрофагами и процедуру окрашивания проводили, как описано в и . f Секреция ТНТ в цитозоль инфицированных макрофагов штаммов Mtb h47Rv и Erdman. Заражение макрофагами и процедуру окрашивания проводили, как описано в и .По показаниям бактерии выращивали в 7H9 + 100 нг/мл ангидротетрациклина (ATc) для предварительного истощения EccD5 перед инфицированием, затем ATc сохраняли в культуральной среде во время инфицирования. Штамм Δ esx-1 имеет делецию области RD1 и лишен генов panCD . Данные представлены как среднее ± SEM по крайней мере двух независимых экспериментов ( n ≥ 2), и показаны репрезентативные изображения. Звездочки указывают на существенные различия (* p значение ≤ 0,05, ** p значение ≤ 0.01, *** p значение ≤ 0,001, **** p значение ≤ 0,0001, рассчитанное с использованием однофакторного дисперсионного анализа с поправкой Даннета) по сравнению со штаммом Mtb mc 2 6206. Исходные данные предоставляются в файле исходных данных.
Звездочки указывают на существенные различия (* p значение ≤ 0,05, ** p значение ≤ 0.01, *** p значение ≤ 0,001, **** p значение ≤ 0,0001, рассчитанное с использованием однофакторного дисперсионного анализа с поправкой Даннета) по сравнению со штаммом Mtb mc 2 6206. Исходные данные предоставляются в файле исходных данных.
Системы ESX-1, ESX-2 и ESX-4 необходимы для TNT-опосредованной цитотоксичности у
M. tuberculosis NAD + гликогидролаза TNT токсична только тогда, когда она имеет доступ к NAD + в клеточном цитозоле 6 .Таким образом, мы предположили, что мутанты Mtb ESX с нарушенной фагосомной проницаемостью обладают сниженной цитотоксичностью в макрофагах из-за нарушения переноса токсинов. Чтобы проверить эту гипотезу, мы измерили жизнеспособность макрофагов THP-1, инфицированных Mtb mc 2 6206 и esx-1 , esx-2, и esx-4 -дефицитных мутантов. Штамм Mtb mc 2 6206 обладает той же цитотоксичностью, что и его вирулентный родительский штамм Mtb h47Rv (дополнительные рисунки 5, 6), что согласуется с предыдущим отчетом 21 .Цитотоксичность штамма Mtb, лишенного оперона cpnT , в макрофагах была в значительной степени снижена (рис. 4d). Комплементация оперона cpnT полностью восстановила его цитотоксичность до уровня дикого типа (рис. 4d) в соответствии с предыдущими результатами, демонстрирующими, что CpnT и, в частности, каталитическая активность его домена TNT, является основным фактором цитотоксичности в макрофагах, инфицированных Mtb . 6, 7 . Важно отметить, что цитотоксичность штаммов Δ esx-1 , Δ esx-2, и Δ esx-4 была снижена до уровня, аналогичного таковому у штамма Δ cpnT (рис.4г). Цитотоксичность была восстановлена, когда соответствующие мутанты Mtb были дополнены космидами I106 и I60 (дополнительная таблица 2), содержащими локусы esx-2 или esx-4 соответственно (рис.
Штамм Mtb mc 2 6206 обладает той же цитотоксичностью, что и его вирулентный родительский штамм Mtb h47Rv (дополнительные рисунки 5, 6), что согласуется с предыдущим отчетом 21 .Цитотоксичность штамма Mtb, лишенного оперона cpnT , в макрофагах была в значительной степени снижена (рис. 4d). Комплементация оперона cpnT полностью восстановила его цитотоксичность до уровня дикого типа (рис. 4d) в соответствии с предыдущими результатами, демонстрирующими, что CpnT и, в частности, каталитическая активность его домена TNT, является основным фактором цитотоксичности в макрофагах, инфицированных Mtb . 6, 7 . Важно отметить, что цитотоксичность штаммов Δ esx-1 , Δ esx-2, и Δ esx-4 была снижена до уровня, аналогичного таковому у штамма Δ cpnT (рис.4г). Цитотоксичность была восстановлена, когда соответствующие мутанты Mtb были дополнены космидами I106 и I60 (дополнительная таблица 2), содержащими локусы esx-2 или esx-4 соответственно (рис. 4d). Эти результаты согласуются с предыдущими экспериментами, согласно которым системы ESX-1, -2 и -4 необходимы для CpnT/TNT для достижения цитозоля макрофагов, инфицированных Mtb.
4d). Эти результаты согласуются с предыдущими экспериментами, согласно которым системы ESX-1, -2 и -4 необходимы для CpnT/TNT для достижения цитозоля макрофагов, инфицированных Mtb.
Системы ESX-1, ESX-2 и ESX-4 необходимы для фагосомной пермеабилизации в
M.туберкулез Основываясь на наблюдении, что доступ TNT к цитозолю хозяина требует системы ESX-2, мы предположили, что система ESX-2 может иметь сходную с ESX-1 функцию в разрыве фагосомы 9, 10 . Чтобы исключить какой-либо CpnT/TNT-специфический эффект, мы исследовали роль систем Mtb ESX в перемещении цитозольных белков с использованием антисыворотки против белков Mtb в производном туберкулинового очищенного белка (PPD) 33 . С этой целью мембраны инфицированных макрофагов селективно пермеабилизировали с использованием дигитонина, как описано выше.Антитела в PPD-антисыворотке обнаруживали белки после обработки дигитонином, что указывает на пермеабилизацию фагосомальной мембраны родительским штаммом Mtb mc 2 6206. Это согласуется с ранее наблюдаемым цитоплазматическим доступом TNT в Mtb. Интересно, что обнаруженные белки, по-видимому, находятся в непосредственной близости от клеток Mtb, в отличие от TNT, который распространяется по цитоплазме инфицированных макрофагов (рис. 5a–c, дополнительная рис. 17). Как и ожидалось, белки Mtb не были обнаружены после обработки дигитонином макрофагов, инфицированных штаммом Mtb, лишенным системы ESX-1 (рис.5a – c, дополнительный рисунок 17). Этот результат согласуется со многими сообщениями о том, что Mtb требует системы ESX-1 для разрыва фагосомы, и нашими предыдущими результатами о том, что ESX-1 необходим для TNT для доступа к цитозолю инфицированных макрофагов. Этот эксперимент также подтверждает, что пермеабилизацию фагосом можно определить путем доступа антител к ассоциированным с клеткой или секретируемым белкам Mtb в дополнение к обнаружению TNT, как показано на рис. 4a. Поразительно, но белки Mtb не были обнаружены в пермеабилизированных дигитонином макрофагах, инфицированных штаммами esx-2- и esx-4- с дефицитом, тогда как оба штамма продуцировали обнаруживаемые белки после пермеабилизации Triton X-100 (рис.
Это согласуется с ранее наблюдаемым цитоплазматическим доступом TNT в Mtb. Интересно, что обнаруженные белки, по-видимому, находятся в непосредственной близости от клеток Mtb, в отличие от TNT, который распространяется по цитоплазме инфицированных макрофагов (рис. 5a–c, дополнительная рис. 17). Как и ожидалось, белки Mtb не были обнаружены после обработки дигитонином макрофагов, инфицированных штаммом Mtb, лишенным системы ESX-1 (рис.5a – c, дополнительный рисунок 17). Этот результат согласуется со многими сообщениями о том, что Mtb требует системы ESX-1 для разрыва фагосомы, и нашими предыдущими результатами о том, что ESX-1 необходим для TNT для доступа к цитозолю инфицированных макрофагов. Этот эксперимент также подтверждает, что пермеабилизацию фагосом можно определить путем доступа антител к ассоциированным с клеткой или секретируемым белкам Mtb в дополнение к обнаружению TNT, как показано на рис. 4a. Поразительно, но белки Mtb не были обнаружены в пермеабилизированных дигитонином макрофагах, инфицированных штаммами esx-2- и esx-4- с дефицитом, тогда как оба штамма продуцировали обнаруживаемые белки после пермеабилизации Triton X-100 (рис. 5a – c, дополнительный рисунок 17). Доступ антител к белкам Mtb в макрофагах, обработанных дигитонином, был восстановлен путем экспрессии локусов esx-2 и esx-4 в соответствующих мутантах Mtb с использованием космид I106 и I60 (дополнительная таблица 2) соответственно (рис. 5a). , б, дополнительный рис. 17). Поскольку уровни белка субстрата ESX-1 CFP-10 идентичны в культуральных фильтратах штаммов esx-2- и esx-4- с дефицитом и его родительского штамма Mtb mc 2 6206 (дополнительная рис.8), мы пришли к выводу, что отсутствие фагосомной пермеабилизации в макрофагах, инфицированных штаммами esx-2- и esx-4-, не связано с нарушением системы ESX-1. Следовательно, эти результаты демонстрируют, что системы ESX-2 и ESX-4 необходимы, в дополнение к ESX-1, для пермеабилизации фагосомальной мембраны и переноса белка между фагосомой и цитозолем макрофагов, инфицированных Mtb. Эти результаты раскрывают первую известную молекулярную функцию системы ESX-2 у Mtb и показывают, что система ESX-4 имеет по крайней мере две функции: экспорт и секреция CpnT/TNT Mtb и пермеабилизация фагосомы вместе с ESX-1.
5a – c, дополнительный рисунок 17). Доступ антител к белкам Mtb в макрофагах, обработанных дигитонином, был восстановлен путем экспрессии локусов esx-2 и esx-4 в соответствующих мутантах Mtb с использованием космид I106 и I60 (дополнительная таблица 2) соответственно (рис. 5a). , б, дополнительный рис. 17). Поскольку уровни белка субстрата ESX-1 CFP-10 идентичны в культуральных фильтратах штаммов esx-2- и esx-4- с дефицитом и его родительского штамма Mtb mc 2 6206 (дополнительная рис.8), мы пришли к выводу, что отсутствие фагосомной пермеабилизации в макрофагах, инфицированных штаммами esx-2- и esx-4-, не связано с нарушением системы ESX-1. Следовательно, эти результаты демонстрируют, что системы ESX-2 и ESX-4 необходимы, в дополнение к ESX-1, для пермеабилизации фагосомальной мембраны и переноса белка между фагосомой и цитозолем макрофагов, инфицированных Mtb. Эти результаты раскрывают первую известную молекулярную функцию системы ESX-2 у Mtb и показывают, что система ESX-4 имеет по крайней мере две функции: экспорт и секреция CpnT/TNT Mtb и пермеабилизация фагосомы вместе с ESX-1. и ESX-2, что обеспечивает диффузию токсина в цитозоль макрофагов.
и ESX-2, что обеспечивает диффузию токсина в цитозоль макрофагов.
a Обнаружение разрыва фагосомы в инфицированных макрофагах с помощью Mtb-специфических антител. Указанные штаммы Mtb были помечены метаболическим красителем DMN-трегалоза (зеленый) и использованы для заражения макрофагов THP-1 при множественности заражения 10:1. Через 48 ч после заражения макрофаги пермеабилизировали дигитонином, чтобы обеспечить доступ антител к цитоплазме, или тритоном Х-100, чтобы получить доступ к внутриклеточным компартментам.Затем клетки окрашивали антителом против Mtb (αMtb) и вторичным антителом Alexa Fluor-594 (красный). Ядра макрофагов окрашивали DAPI. b , c Количественное определение αMtb-положительных макрофагов после пермеабилизации дигитонином ( b ) и Triton X-100 ( c ) из изображений, показанных в a . Макрофаги оценивали как αMtb-положительные, когда наблюдались отчетливые красные точки по сравнению с мутантом Mtb esx-1 , обработанным дигитонином. d Обнаружение разорванных фагосом в макрофагах, инфицированных Mtb. Указанные штаммы Mtb были помечены метаболическим красителем DMN-трегалоза (зеленый) и использованы для заражения макрофагов THP-1 при множественности заражения 10:1. Через 48 ч после заражения макрофаги пермеабилизировали с помощью Triton X-100 и окрашивали антителами против маркера разрыва фагосомы галектин-3 и вторичными антителами Alexa Fluor-594 (красный). и . Количественное определение галектин-3-положительных макрофагов по изображениям, показанным в d .Макрофаги считали положительными по галектину-3, когда наблюдали отчетливые красные точки по сравнению с мутантом Mtb esx-1 . Штамм Δ esx-1 имеет делецию области RD1 и лишен генов panCD . Данные представлены как среднее ± SEM по крайней мере двух независимых экспериментов ( n ≥ 2), и показаны репрезентативные изображения.
Макрофаги оценивали как αMtb-положительные, когда наблюдались отчетливые красные точки по сравнению с мутантом Mtb esx-1 , обработанным дигитонином. d Обнаружение разорванных фагосом в макрофагах, инфицированных Mtb. Указанные штаммы Mtb были помечены метаболическим красителем DMN-трегалоза (зеленый) и использованы для заражения макрофагов THP-1 при множественности заражения 10:1. Через 48 ч после заражения макрофаги пермеабилизировали с помощью Triton X-100 и окрашивали антителами против маркера разрыва фагосомы галектин-3 и вторичными антителами Alexa Fluor-594 (красный). и . Количественное определение галектин-3-положительных макрофагов по изображениям, показанным в d .Макрофаги считали положительными по галектину-3, когда наблюдали отчетливые красные точки по сравнению с мутантом Mtb esx-1 . Штамм Δ esx-1 имеет делецию области RD1 и лишен генов panCD . Данные представлены как среднее ± SEM по крайней мере двух независимых экспериментов ( n ≥ 2), и показаны репрезентативные изображения. Звездочки указывают на существенные различия (* p значение ≤ 0,05, ** p значение ≤ 0,01, *** p значение ≤ 0.001, **** p значение ≤ 0,0001, рассчитанное с использованием однофакторного дисперсионного анализа с поправкой Даннета) по сравнению со штаммом Mtb Δ esx-1 . Исходные данные предоставляются в файле исходных данных.
Звездочки указывают на существенные различия (* p значение ≤ 0,05, ** p значение ≤ 0,01, *** p значение ≤ 0.001, **** p значение ≤ 0,0001, рассчитанное с использованием однофакторного дисперсионного анализа с поправкой Даннета) по сравнению со штаммом Mtb Δ esx-1 . Исходные данные предоставляются в файле исходных данных.
Мы подозревали, что системы ESX-2 и ESX-4 способствуют разрыву фагосомальной мембраны за счет мембранолитической активности, сходным образом с ESX-1 34, 35 . Чтобы проверить эту гипотезу, мы использовали galectin-3, белок-хозяин, который распознает люменальные гликаны, которые подвергаются воздействию цитозоля, когда фагосома сильно повреждена 36,37,38 .Действительно, макрофаги, инфицированные Mtb, показали значительное повреждение мембран рядом с бактериями, как визуализировали с помощью антитела к галектину-3 (рис. 5d, e, дополнительная рис. 18). Как и ожидалось, окрашивание галектин-3 отсутствовало в макрофагах, инфицированных штаммом Δ esx-1 , и сильно снижалось в макрофагах, инфицированных штаммами Δ esx-2 и Δ esx-4 (рис. 5г, д, Дополнительный рис. 18). Важно отметить, что окрашивание галектина-3 было восстановлено до уровней дикого типа путем экспрессии локусов esx-2 и esx-4 с использованием космид I106 и I60 (дополнительная таблица 2) в соответствующих мутантах Mtb (рис.5d, e, дополнительный рисунок 18), демонстрирующий, что системы ESX-2 и ESX-4 действительно повреждают фагосому, содержащую Mtb. Количественная оценка окрашивания галектином показала, что система ESX-1 вызывает большее повреждение мембран по сравнению с системой ESX-2 (рис. 5e). Напротив, эксперименты по пермеабилизации с использованием антител TNT и Mtb показали идентичные фенотипы для штаммов с дефицитом esx-1 , esx-2, и esx-4 . Следовательно, мы заключаем, что для пермеабилизации фагосом необходима скоординированная активность всех трех систем ESX.В совокупности эти результаты показывают, что система ESX-4 не только необходима для секреции TNT Mtb, но также играет важную роль в проницаемости фагосомной мембраны вместе с ESX-1 и ESX-2, чтобы обеспечить перенос TNT и других веществ.
5г, д, Дополнительный рис. 18). Важно отметить, что окрашивание галектина-3 было восстановлено до уровней дикого типа путем экспрессии локусов esx-2 и esx-4 с использованием космид I106 и I60 (дополнительная таблица 2) в соответствующих мутантах Mtb (рис.5d, e, дополнительный рисунок 18), демонстрирующий, что системы ESX-2 и ESX-4 действительно повреждают фагосому, содержащую Mtb. Количественная оценка окрашивания галектином показала, что система ESX-1 вызывает большее повреждение мембран по сравнению с системой ESX-2 (рис. 5e). Напротив, эксперименты по пермеабилизации с использованием антител TNT и Mtb показали идентичные фенотипы для штаммов с дефицитом esx-1 , esx-2, и esx-4 . Следовательно, мы заключаем, что для пермеабилизации фагосом необходима скоординированная активность всех трех систем ESX.В совокупности эти результаты показывают, что система ESX-4 не только необходима для секреции TNT Mtb, но также играет важную роль в проницаемости фагосомной мембраны вместе с ESX-1 и ESX-2, чтобы обеспечить перенос TNT и других веществ. белков в цитозоль макрофагов, инфицированных Mtb.
белков в цитозоль макрофагов, инфицированных Mtb.
Познакомьтесь с Ольгой Фондой, которую мы скромно номинируем на роль будущей девушки Бонда
Как можно начать описывать Ольгу Фонду? Женщины хотят быть ею, мужчины хотят быть с ней, и никто не хочет перечить ей; она занимается карате и крав-магой, и, вероятно, могла бы покалечить тебя и при этом хорошо выглядеть.В Agent X Фонда играет русского шпиона, за которым охотится безжалостный агент, титульный агент X, в то время как вице-президент Шэрон Стоун председательствует, и она уже доказывает, что у нее есть все разнообразие женских отбивных Бонда.
Фонда переехала в Соединенные Штаты на длительный срок вскоре после получения диплома по бизнесу в одном из московских университетов. Ее обнаружили в Лос-Анджелесе, и она начала работать моделью, а вскоре и актерской. Впервые мы обратили внимание на 33-летнюю русскую, когда она появилась в сериале «Дневники вампира», эпизод , который мы могли со стыдом смотреть все семь сезонов, как умного, опасного вампира с сексуальным акцентом. Нам нужно было узнать больше, поэтому мы позвонили ей в Лос-Анджелес, чтобы узнать, чем она занимается (бразильская подтяжка ягодиц), и насладиться нашей общей любовью к Шэрон Стоун.
Нам нужно было узнать больше, поэтому мы позвонили ей в Лос-Анджелес, чтобы узнать, чем она занимается (бразильская подтяжка ягодиц), и насладиться нашей общей любовью к Шэрон Стоун.
GQ: Где мы с вами находимся?
Ольга Фонда: Вообще-то я звоню тебе из машины, но я в парке! Я переехал примерно в 45 минутах езды от Западного Голливуда и живу в окружении природы и дикой природы, но я постоянно ловлю себя на том, что хожу, как в рекламе, и говорю: «Теперь ты меня слышишь?» Пришлось объехать окрестности, чтобы найти место.
Я читал, что вы выбрали Фонда в качестве своей фамилии, когда переехали в Соединенные Штаты. Почему Фонда?
Вопрос на миллион долларов! Я хотел бы сохранить это в тайне.
Как вы проводите свободное время между проектами?
Как и все, я стараюсь выполнять все свои поручения. Я стараюсь видеться с друзьями и семьей. Несколько недель назад я пошел и впервые увидел Звуки музыки . Он играл в центре Лос-Анджелеса, и мне это очень понравилось.Я стираю белье, постоянно работаю над улучшением своих актерских навыков и стараюсь оставаться в форме.
Он играл в центре Лос-Анджелеса, и мне это очень понравилось.Я стираю белье, постоянно работаю над улучшением своих актерских навыков и стараюсь оставаться в форме.
Как вы это делаете? Прошу друга.
Я хожу в школу карате рядом с моим домом!
Что тебе нравится в каратэ?
Я думаю, что это восходит к детству. Долгое время мой старший брат говорил мне, что учит меня самообороне, но теперь, когда я вырос, я понимаю, что он просто практиковал на мне свои боевые искусства. Я также только что обнаружил DVD с домашними упражнениями — не смейтесь — он называется «Бразильская подтяжка ягодиц».Это очень увлекательная тренировка, которую я делаю сама. Если вы никогда не пробовали это, это очень весело. Много танцев. Из трехмесячного испытания у меня вторая неделя.
«Дверь открылась, и это было как в замедленной съемке. Эта красивая женщина входит — дует ветер, вся комната просто замирает. Я почувствовал, как мое сердце буквально екнуло.
 » — на встречу Шэрон Стоун
» — на встречу Шэрон Стоун Удачи с оставшейся десяткой!
Это весело. Мой муж такой забавный; Я закрываю дверь и говорю, что мне нужно уединение, чтобы танцевать.Но он постоянно забывает свои ключи или телефон, и я знаю, что втайне он просто хочет увидеть меня танцующей в моем милом наряде.
Уверен, наши читатели поступили бы так же. Расскажите мне о своем персонаже в Agent X .
Каждый мужчина хочет быть с ней, и каждая девушка хочет быть ее другом. Она очень захватывающая, любит опасность и обладает отличными боевыми навыками.
Боевые навыки! Удалось ли вам узнать что-то новое для шоу?
Да.Мы снимали в Ванкувере, и я приехал на несколько недель раньше и работал в студии крав-мага, осваивая новые навыки. Было очень весело, и я старался делать столько своих собственных трюков, сколько мне позволяли. У нас отличная команда каскадеров, но я действительно люблю делать это сам, когда у меня есть шанс.
TNT выпускает сериал «Сквозь снег»?
Серия popSnowpiercer выходит на телевидение
Согласно отчету Collider, TNT в настоящее время работает над созданием телесериала по хиту Sleeper Snowpiercer .
Для тех из вас, кто забыл посмотреть этот невероятно оригинальный фильм, вот синопсис.
«Прошло 18 лет с тех пор, как мы заморозили землю. Несколько оставшихся людей живут на «Сквозь снег», поезде, который движется по бесконечной петле вокруг земного шара. Для тех, кто на передовой, это роскошный рай с наркотиками и суши на лоне роскоши; для тех, кто попал в хвостовую часть, жизнь коротка и жестока.
Но перемены витают в воздухе. Кертис (Крис Эванс), отчаянно пытающийся вырваться из хвоста поезда, планирует восстание, которому помогает его наставник Гиллиам (Джон Хёрт).То, что начинается как изолированный бунт, перерастает в массовую революцию, толчок вперед по принципу «все или ничего» и войну за будущее человечества.
 Кто будет жить, а кто умрет? Как далеко они могут зайти? Есть ли надежда за пределами замороженных пустошей? (Официальный сайт Snowpiercer)
Кто будет жить, а кто умрет? Как далеко они могут зайти? Есть ли надежда за пределами замороженных пустошей? (Официальный сайт Snowpiercer)Зачем вам это?
С появлением всех фильмов и телевизионных шоу вполне логично, что вы могли пропустить этот фильм. «Сквозь снег » — один из самых оригинальных и креативных фильмов 2014 года.Научно-фантастический фильм корейского режиссера Бонг Джун-Хо мрачный и неуклонно говорит о неравенстве между социальными классами.
2016 год кажется эскалацией в этих темах, поскольку новости пролили свет на многие несправедливости, происходящие в США. Кажется уместным, что мы хотели бы исследовать это дальше.
Snowpiercer успешно строит завораживающий мир, в котором многие хотят большего. История этой вымышленной земли созрела для исследования. Действие сериала TNT будет происходить через 7 лет после того, как мир замерзнет, как и в фильме.
Как это будет работать?
Если вы видели Snowpiercer , вы можете удивиться, как можно построить целую серию из нескольких вагонов поезда. Однако фильм основан на французском графическом романе Le Transperceneige Жака Лоба и Жан-Марка Рошета. В романе поезд состоит из 1001 вагона, что действительно открывает серию для исследования каждого закоулка.
Однако фильм основан на французском графическом романе Le Transperceneige Жака Лоба и Жан-Марка Рошета. В романе поезд состоит из 1001 вагона, что действительно открывает серию для исследования каждого закоулка.
Серия Snowpiercer действительно может расширить первоначальный мир, созданный по фильму.Я действительно с нетерпением жду этого, но я буду очень скучать по Тильде Суинтон.
Следите за новостями в блоге! Вот трейлер для Snowpiercer ! Смотри.
Брайан Майя
SENTINELLE Трейлер: Французский солдат Ольга Куриленко ищет мести за свою сестру в фильме Жюльена Леклерка 2020 года
Трейлер Сентинелл
Julien Leclercq s Sentinelle ( 2020 ) Трейлер фильма был выпущен Netflix . Sentinelle Trailer Sentinelle oLga Kurylenko , Andrey Gorlenko , Gabriel Afonso , Gabriel ALMAER , Michel Biel , Julian de Backer , Guulian de Backer , Guulaume Duese , IDRIS IBRAGIMOV , Temerlan Idigov , Мэрилин Лима , Антония Малинова , Мишель Набокофф , Мартин Сваби и Кэрол Вейерс .
Экипаж
Жюльен Леклерк и Матье Серво написали сценарий для Сентинель . Солин Гийонно вела монтаж этого фильма. Брехт Гойвертс создал операторскую работу для фильма.
Реклама
Краткий обзор сюжета
Сюжет Sentinelle : «Кларе тридцать три года. Она переводчик во французской армии. Пережив ужасы войны в Сирии, она была переведена в Ниццу в рамках операции «Страж». Там с матерью и сестрой Таней она пытается восстановить себя.Но однажды вечером, после похода в ночной клуб, Таню находят полумертвой на пляже. Ее изнасиловали и избили. Затем Клара сделает все, чтобы найти нападавших и отомстить за свою сестру. Эта беспощадная охота приведет ее по стопам Ивана Кадникова, сына влиятельного русского олигарха Французской Ривьеры».
Мысли
Выглядит хорошо, а сцены драк кажутся довольно жестокими. Куриленко фактически продает себя французским солдатом. Это может помочь, потому что я не видел ее в течение некоторого времени.
Смотреть трейлер Sentinel . Оставьте свои мысли по этому поводу ниже в разделе комментариев. Читатели, желающие поддержать этот тип контента, могут посетить нашу страницу Patreon и стать одним из покровителей FilmBook. Читатели, которые ищут больше трейлеров к фильмам, могут посетить нашу страницу с трейлерами к фильмам, нашу страницу с трейлерами к фильмам в Twitter и нашу страницу с трейлерами к фильмам на Facebook. Sentinelle выйдет на Netflix 5 марта 2021 года. Хотите получать самые свежие уведомления? Сотрудники FilmBook публикуют статьи по электронной почте, Twitter, Facebook, Instagram, Tumblr, Pinterest, Reddit и Flipboard.
Сентинель Трейлер 2
ТНТ, цитонемы или межклеточные мостики?
ВВЕДЕНИЕ [1]. Эти структуры, по-видимому, переносят везикулы между соединенными клетками и были названы туннельными нанотрубками (ТНТ), чтобы отличить их от других клеточных выступов и за их уникальное свойство обеспечивать бесшовную непрерывность между цитозолями соединенных клеток, как это делают туннели. ТНТ парили над субстратом и соединяли клетки на больших расстояниях. Поразительно, хотя ТНТ могли достигать длины в несколько диаметров клетки, не прикрепляясь к субстрату, они, по-видимому, не содержали микротрубочек, а только актиновые филаменты. Основываясь на своих данных, авторы предположили, что ключевой характеристикой ТНТ является их способность устанавливать цитоплазматическую непрерывность, что, в свою очередь, позволяет перемещать мембранные везикулы между соединенными клетками [1]. Давно было известно, что клетки образуют информационные сети, контактируя с соседними клетками, а также выпуская диффузных мессенджеров, которые распространяются на большие расстояния, но возможность того, что клетки могут открываться друг другу (пусть даже временно) для прямой связи на большие расстояния не принималось во внимание, поскольку противоречило догме о клетках как об отдельных объектах.Действительно, гипотеза была воспринята научным сообществом с некоторым скептицизмом [2].
ТНТ парили над субстратом и соединяли клетки на больших расстояниях. Поразительно, хотя ТНТ могли достигать длины в несколько диаметров клетки, не прикрепляясь к субстрату, они, по-видимому, не содержали микротрубочек, а только актиновые филаменты. Основываясь на своих данных, авторы предположили, что ключевой характеристикой ТНТ является их способность устанавливать цитоплазматическую непрерывность, что, в свою очередь, позволяет перемещать мембранные везикулы между соединенными клетками [1]. Давно было известно, что клетки образуют информационные сети, контактируя с соседними клетками, а также выпуская диффузных мессенджеров, которые распространяются на большие расстояния, но возможность того, что клетки могут открываться друг другу (пусть даже временно) для прямой связи на большие расстояния не принималось во внимание, поскольку противоречило догме о клетках как об отдельных объектах.Действительно, гипотеза была воспринята научным сообществом с некоторым скептицизмом [2].–
Однако в течение последних 15 лет TNT-подобные структуры наблюдались в различных клеточных линиях в культуре, в первичных клетках и в тканях, и было показано, что они обеспечивают перенос различных грузов, от ионов до органелл [обзор в [3]. ][4][5][6][7]]. На основе наблюдений за перемещением различных грузов предложено несколько функций тротилов. Среди них было показано, что TNT способны электрически соединять клетки на больших расстояниях, обеспечивая передачу сигналов Ca 2+ между соединенными клетками через щелевые контакты или рецепторы IP 3 [3][8].Это может согласовываться с участием TNT в развитии, когда электрическая связь может помочь организовать морфогенетические движения, которые позволяют клеткам достигать своей специфической ниши, где будет происходить дифференцировка в различных тканях или типах клеток [9][10]. В частности, электрическая связь через TNT может иметь решающее значение во время раннего развития мозга, когда нейронные предшественники еще не установили зрелые химические синапсы [3][4][5][11]. Перенос кальция через ТНТ, установленный между миелоидными клетками, также предполагает участие ТНТ в активации иммунных клеток и иммунном ответе [8][12].Участие TNT в иммунном ответе также подтверждается наблюдением TNT-опосредованного переноса Fas-лиганда сигнала смерти и активных каспаз, что приводит к индукции апоптоза в принимающих клетках [13].
][4][5][6][7]]. На основе наблюдений за перемещением различных грузов предложено несколько функций тротилов. Среди них было показано, что TNT способны электрически соединять клетки на больших расстояниях, обеспечивая передачу сигналов Ca 2+ между соединенными клетками через щелевые контакты или рецепторы IP 3 [3][8].Это может согласовываться с участием TNT в развитии, когда электрическая связь может помочь организовать морфогенетические движения, которые позволяют клеткам достигать своей специфической ниши, где будет происходить дифференцировка в различных тканях или типах клеток [9][10]. В частности, электрическая связь через TNT может иметь решающее значение во время раннего развития мозга, когда нейронные предшественники еще не установили зрелые химические синапсы [3][4][5][11]. Перенос кальция через ТНТ, установленный между миелоидными клетками, также предполагает участие ТНТ в активации иммунных клеток и иммунном ответе [8][12].Участие TNT в иммунном ответе также подтверждается наблюдением TNT-опосредованного переноса Fas-лиганда сигнала смерти и активных каспаз, что приводит к индукции апоптоза в принимающих клетках [13].
–
И последнее, но не менее важное: считается, что ТНТ способствуют развитию различных патологических состояний. Точно так же способность ТНТ образовываться между опухолевыми клетками и соединять опухолевые клетки со стромальными клетками, обеспечивая перенос микро-РНК и митохондрий между связанными клетками, предполагает участие ТНТ в прогрессировании рака [7][14][15]. [16].Было показано, что ТНТ-подобные структуры переносят ВИЧ-1 между Т-клетками и между макрофагами [17][18], что предполагает участие ТНТ-подобных структур в распространении ВИЧ-1. Также было показано, что прионы и прионоподобные белки используют TNT для своего распространения. В частности, было показано, что ТНТ переносят агрегаты PrP Sc , альфа-синуклеина, тау, бета-амилоида, полиглутамина хантингтина и разрушенного при шизофрении 1 (DISC1) [19, 20, 21, 22, 23]. . Поэтому было высказано предположение, что, позволяя прионоподобным белкам распространяться в головном мозге, ТНТ могут играть заметную роль в патогенезе различных нейродегенеративных заболеваний [22].
–
Одна из основных проблем в области TNT заключается в том, что было описано множество различных TNT-подобных структур в разных типах клеток и в разных условиях [рассмотрено в [24][25][26][27][28]]. Поскольку в этой области все еще отсутствуют маркеры, специфичные для TNT, до сих пор неизвестно, существуют ли один или несколько типов bona fide TNT (открытые структуры, обеспечивающие прямую связь между клетками), и существует ли структурно-функциональная связь между ними. различные структуры.Хотя ясно, что TNT отличаются от других закрытых и более коротких клеточных выступов, таких как реснички и прикрепленные к субстрату филоподии [29][30], все еще проблематично дифференцировать их от подобных структур, соединяющих отдаленные клетки, таких как цитонемы и межклеточные мостики.
–
Принимая во внимание недавнюю идентификацию уникальной структуры ТНТ, образующихся между нервными клетками в культуре [31], в этом обзоре мы специально сосредоточимся на различиях между ТНТ и другими межклеточными структурами (например,грамм. межклеточные мостики и цитонемы) как со структурной, так и с функциональной точки зрения.
межклеточные мостики и цитонемы) как со структурной, так и с функциональной точки зрения.
Возможное участие ТНТ в патогенезе таких заболеваний, как рак, СПИД и нейродегенеративные заболевания, послужило поводом для проведения растущего числа исследований, направленных на характеристику ТНТ. Однако исследования ТНТ сталкиваются с методологическими трудностями из-за хрупкости ТНТ, а именно их чувствительности к химической фиксации, механическим воздействиям и длительному световому возбуждению [1][32].Другой проблемой является отсутствие ТНТ-специфических маркеров, что затрудняет идентификацию ТНТ и изучение их функций в тканях. В связи с этим морфологические свойства остаются главными критериями идентификации ТНТ. Свойства, обычно используемые для идентификации TNT, включают: прямую мостовидную структуру на основе F-актина, соединяющие пары клеток; длина на несколько диаметров клеток и толщина менее 1 мкм. Однако диаметр и длина тротила сильно различаются. Было показано, что длины ТНТ могут меняться по мере того, как связанные клетки раздвигаются или мигрируют, и, следовательно, расстояния между ними меняются.Диаметр ТНТ обычно находится в пределах от 50 до 700 нм, что также зависит от метода, используемого для идентификации ТНТ [рассмотрено в [26][33]]. Кроме того, было показано, что в определенных условиях ТНТ также содержат микротрубочки. Таким образом, было показано, что апоптотические клетки PC12 образуют TNT, содержащие микротрубочки, в то время как TNT тех же клеток PC12, образующиеся в нормальных условиях, лишены микротрубочек [34]. TNT-подобные структуры, генерируемые Т-клетками, которые обеспечивают перенос белка ВИЧ-1 на большие расстояния, по-видимому, содержат микротрубочки.Однако неясно, являются ли они каноническими открытыми ТНТ или закрытыми выступами (см. ниже) [17]. Существует предложение классифицировать ТНТ по их диаметру [35], где «тонкие» нанотрубки имеют диаметр до нескольких сотен нанометров, а «толстые» нанотрубки имеют диаметр более нескольких сотен нанометров.
Было показано, что длины ТНТ могут меняться по мере того, как связанные клетки раздвигаются или мигрируют, и, следовательно, расстояния между ними меняются.Диаметр ТНТ обычно находится в пределах от 50 до 700 нм, что также зависит от метода, используемого для идентификации ТНТ [рассмотрено в [26][33]]. Кроме того, было показано, что в определенных условиях ТНТ также содержат микротрубочки. Таким образом, было показано, что апоптотические клетки PC12 образуют TNT, содержащие микротрубочки, в то время как TNT тех же клеток PC12, образующиеся в нормальных условиях, лишены микротрубочек [34]. TNT-подобные структуры, генерируемые Т-клетками, которые обеспечивают перенос белка ВИЧ-1 на большие расстояния, по-видимому, содержат микротрубочки.Однако неясно, являются ли они каноническими открытыми ТНТ или закрытыми выступами (см. ниже) [17]. Существует предложение классифицировать ТНТ по их диаметру [35], где «тонкие» нанотрубки имеют диаметр до нескольких сотен нанометров, а «толстые» нанотрубки имеют диаметр более нескольких сотен нанометров. Также предполагалось, что «тонкие» нанотрубки могут заканчиваться щелевыми контактами и обеспечивать обмен меньшими грузами, такими как молекулы с молекулярной массой менее 1,2 кДа, включая вторичные мессенджеры и небольшие пептиды, в то время как «толстые» ТНТ могут содержать микротрубочки и, таким образом, быть более стабильными и быть в состоянии мобилизовать более крупные грузы, такие как органеллы или вирусы [33][35].Однако эти критерии недостаточно строги и применимы не ко всем типам клеток.
Также предполагалось, что «тонкие» нанотрубки могут заканчиваться щелевыми контактами и обеспечивать обмен меньшими грузами, такими как молекулы с молекулярной массой менее 1,2 кДа, включая вторичные мессенджеры и небольшие пептиды, в то время как «толстые» ТНТ могут содержать микротрубочки и, таким образом, быть более стабильными и быть в состоянии мобилизовать более крупные грузы, такие как органеллы или вирусы [33][35].Однако эти критерии недостаточно строги и применимы не ко всем типам клеток.
–
Недавнее использование корреляционной световой и криоэлектронной микроскопии предоставило несколько структурных деталей о морфологии ТНТ, которые предполагают, что термины «тонкие» и «толстые» ТНТ следует использовать с осторожностью [31]. В этом исследовании TNT были сохранены ближе к своему нативному статусу благодаря фиксации быстрым замораживанием. Это позволило получить изображения с нанометровым разрешением с помощью коррелятивной криофлуоресцентной и криоэлектронной микроскопии и томографии в условиях полной гидратации, которые являются наилучшими для сохранения мембранных структур. Полученные данные показывают, что структуры, которые при флуоресцентной микроскопии кажутся более толстыми ТНТ, на самом деле могут состоять из нескольких отдельных туннелирующих нанотрубок (названных iTNT) (рис. 1) . Каждый iTNT содержал пучки актина, которые в большинстве случаев заполняли весь просвет трубки. Средний диаметр iTNT около 120 нм и такая плотная упаковка действующих пучков, по-видимому, не препятствовали iTNT переносу везикулярных компартментов и даже митохондрий внутри их просвета, поскольку мембрана часто выпячивалась, чтобы приспособиться к прохождению везикулы (Рисунок 1) .Кроме того, криоэлектронная томография показала наличие тонких коротких филаментов, меченных антителами к N-кадгерину, которые, по-видимому, соединяют iTNT между собой, возможно, для удержания их в пучке и придания более высокой механической стабильности [31] (рис. 1) . В дополнение к пучкам iTNT также наблюдались одиночные более толстые TNT (диаметром 600-900 нм).
Полученные данные показывают, что структуры, которые при флуоресцентной микроскопии кажутся более толстыми ТНТ, на самом деле могут состоять из нескольких отдельных туннелирующих нанотрубок (названных iTNT) (рис. 1) . Каждый iTNT содержал пучки актина, которые в большинстве случаев заполняли весь просвет трубки. Средний диаметр iTNT около 120 нм и такая плотная упаковка действующих пучков, по-видимому, не препятствовали iTNT переносу везикулярных компартментов и даже митохондрий внутри их просвета, поскольку мембрана часто выпячивалась, чтобы приспособиться к прохождению везикулы (Рисунок 1) .Кроме того, криоэлектронная томография показала наличие тонких коротких филаментов, меченных антителами к N-кадгерину, которые, по-видимому, соединяют iTNT между собой, возможно, для удержания их в пучке и придания более высокой механической стабильности [31] (рис. 1) . В дополнение к пучкам iTNT также наблюдались одиночные более толстые TNT (диаметром 600-900 нм). Важно отметить, что пучки iTNT и более толстые одиночные TNT нельзя было различить с помощью флуоресцентной микроскопии, поскольку они имели схожий внешний вид.Эти данные свидетельствуют о том, что может быть неточным применение классификации ТНТ на основе диаметра для идентификации ТНТ с помощью флуоресцентной микроскопии, поскольку «толстые» ТНТ могут в равной степени представлять собой пучки иТНТ, которые изначально имеют тот же диаметр, что и «тонкие» ТНТ.
Важно отметить, что пучки iTNT и более толстые одиночные TNT нельзя было различить с помощью флуоресцентной микроскопии, поскольку они имели схожий внешний вид.Эти данные свидетельствуют о том, что может быть неточным применение классификации ТНТ на основе диаметра для идентификации ТНТ с помощью флуоресцентной микроскопии, поскольку «толстые» ТНТ могут в равной степени представлять собой пучки иТНТ, которые изначально имеют тот же диаметр, что и «тонкие» ТНТ.
–
РИСУНОК 1: «Толстые» и «тонкие» соединения TNT. Криоэлектронная микроскопия показывает, что ТНТ могут быть либо одиночным толстым соединением, либо пучком тонких отдельных ТНТ (iTNT).Внутри пучка могут присутствовать как открытые, так и закрытые выступы. Каждый iTNT содержит пучки актина, может содержать везикулы и митохондрии. Тонкие и короткие мембранные нити соединяют несколько iTNT, которые, по-видимому, растут в противоположных направлениях. Адаптировано из Sartori-Rupp et al. |
 [31].
[31].–
Более того, криоэлектронная томография в нейрональных клетках показала, что как тонкие, так и более толстые iTNT содержат только актин, а не микротрубочки [31], в отличие от того, что было предложено в качестве отличительного признака толстых TNT в макрофагах [35].Особый интерес представляет то, что митохондрии, как было показано, транспортируются внутри iTNTs, предположительно, на актиновых рельсах по тубулин-независимому механизму. Это говорит о том, что TNT не обязательно нуждаются в микротрубочках, чтобы обеспечить перенос более крупных грузов, таких как везикулы и органеллы [31].
–
В целом мы предполагаем, что такие морфологические параметры, как длина, толщина и протяженность между двумя клетками, являются недостаточными характеристиками для идентификации ТНТ. С другой стороны, как первоначально было предложено Гердесом и его коллегами, характерными особенностями TNT, которые следует учитывать для определения TNT, являются: 1) установление непрерывности цитоплазмы и 2) возможность переноса различных грузов, включая органеллы, такие как лизосомы, эндосомы и митохондрии.
–
Несмотря на то, что непрерывность цитоплазмы была указана в качестве основного критерия определения ТНТ, четко продемонстрировать ее было сложно. Мы специально рассмотрели вопрос, действительно ли TNT, которые мы наблюдали в клеточных линиях нейронов CAD и SHSY5Y, соединяют цитозоль двух клеток. С этой целью мы выполнили сканирующую электронную микроскопию с фокусированным ионным пучком (FIB-SEM), которая позволила нам отобразить концы (или места контакта) ТНТ. Интересно, что мы смогли идентифицировать как открытые, так и закрытые соединения, такие как инвагинации в контактных зонах (рис. 1) .Мы не могли различить, были ли наблюдаемые различия результатом существования различных типов TNT или они были вызваны временными событиями до или после слияния. Тем не менее, эти данные представляют собой первое прямое подтверждение того, что TNT с открытым концом существуют и могут соответствовать функциональным структурам TNT, наблюдаемым с помощью флуоресцентной микроскопии [31]. Интересно, что мы не могли наблюдать наличие комплексов щелевых контактов на концах тонких iTNT по сравнению с толстыми, в то время как они наблюдались в других типах клеток [27][36].В будущем может быть интересно провести дальнейшее исследование структур TNT, чтобы понять, является ли присутствие коннексинов на кончиках TNT специфичным для типа клеток и/или может быть использовано для определения специфического типа TNT.
Интересно, что мы не могли наблюдать наличие комплексов щелевых контактов на концах тонких iTNT по сравнению с толстыми, в то время как они наблюдались в других типах клеток [27][36].В будущем может быть интересно провести дальнейшее исследование структур TNT, чтобы понять, является ли присутствие коннексинов на кончиках TNT специфичным для типа клеток и/или может быть использовано для определения специфического типа TNT.
–
Анализ криоэлектронной томографии также позволил выявить морфологические различия между филоподиями и ТНТ в нейрональных клетках. Филоподии в нейрональных клетках представляют собой изолированные выпячивания с близким к ТНТ средним диаметром 200 нм. По сравнению с ТНТ филоподии имеют более короткий диапазон длины, от одного до нескольких микрон, и, по-видимому, не содержат пузырьков.Интересно, что филоподии также демонстрируют различное расположение актина по сравнению с TNT, а именно либо плотные параллельные пучки, но меньшей длины по сравнению с TNT, либо параллельные пучки, смешанные с короткоразветвленными филаментами, которые не наблюдались в iTNT. Однако при наблюдении поперечных срезов филоподий было показано, что актиновые филаменты расположены в виде гексагонального массива, подобного наблюдаемому в iTNTs [31].
Однако при наблюдении поперечных срезов филоподий было показано, что актиновые филаменты расположены в виде гексагонального массива, подобного наблюдаемому в iTNTs [31].
–
Тот факт, что TNT обнаруживают некоторые отличия от филоподий в расположении актина, позволяет предположить, что TNT могли формироваться как отдельные структуры с самого начала, а не из филоподий с последующим слиянием их кончика с клеткой-реципиентом.Предыдущие данные о молекулах, регулирующих TNT, подтверждают эту гипотезу. В самом деле, регуляторные молекулы актина могут оказывать противоположное влияние на образование ТНТ и филоподий. В частности, филоподии, индуцирующие сеть CDC42/IRSp53/VASP, уменьшали количество клеток, связанных с TNT, и подавляли опосредованный TNT перенос везикул [30]. Сходным образом, фасцин, для которого ранее было показано, что он индуцирует дорсальные филоподии в нейрональных клетках CAD мыши [37], не способствует образованию TNT в тех же клетках [38]. Напротив, Eps8 (субстрат 8 пути рецептора EGF), другой регулятор актина, который, как было показано, ингибирует образование филоподий в нейронах, оказался положительным регулятором образования TNT и переноса везикул в клетках CAD [30]. Кроме того, наши неопубликованные данные показывают, что Eps8 увеличивает образование ТНТ и перенос α-синуклеина в нейрональные клетки SHSY5Y человека, что позволяет предположить, что он может быть общей молекулой активатора ТНТ для нейронных клеток. Однако следует отметить, что ТНТ и филоподии также могут иметь общие регуляторы своего образования. Аналогично, Myosin X активирует образование как дорсальных филоподий, так и TNT в нейрональных клетках CAD [38]. Следовательно, не следует исключать возможность того, что ТНТ могут возникать из дорсальных филоподий.
Кроме того, наши неопубликованные данные показывают, что Eps8 увеличивает образование ТНТ и перенос α-синуклеина в нейрональные клетки SHSY5Y человека, что позволяет предположить, что он может быть общей молекулой активатора ТНТ для нейронных клеток. Однако следует отметить, что ТНТ и филоподии также могут иметь общие регуляторы своего образования. Аналогично, Myosin X активирует образование как дорсальных филоподий, так и TNT в нейрональных клетках CAD [38]. Следовательно, не следует исключать возможность того, что ТНТ могут возникать из дорсальных филоподий.
–
В целом, мы предполагаем, что морфологические параметры, определенные с помощью световой микроскопии, являются недостаточными характеристиками для идентификации ТНТ, и исследования ТНТ всегда должны быть в состоянии оценить их функциональность в переносе молекул или органелл между связанными клетками. С другой стороны, ТНТ имеют отчетливую структуру, которую можно идентифицировать с помощью электронной микроскопии. Тем не менее, следует отметить, что существует другой тип выпячиваний, которые разделяют с TNT способность обеспечивать непрерывность цитоплазмы и перенос грузов.Это межклеточные мосты (IB), которые, как известно, соединяют клетки зародышевой линии [39][40]. В следующем разделе мы рассмотрим сходства и различия между IB и TNT.
Тем не менее, следует отметить, что существует другой тип выпячиваний, которые разделяют с TNT способность обеспечивать непрерывность цитоплазмы и перенос грузов.Это межклеточные мосты (IB), которые, как известно, соединяют клетки зародышевой линии [39][40]. В следующем разделе мы рассмотрим сходства и различия между IB и TNT.
Межклеточные мостики (IBs) представляют собой цитоплазматические мостики, которые образуются в результате незавершенного цитокинеза и соединяют клетки в синцитиях. Стабильные IB обнаруживаются в женской и мужской зародышевой линии различных организмов, от насекомых до человека. Это эволюционное сохранение предполагает, что эта взаимосвязь зародышевых клеток важна, и действительно было показано, что стабильные межклеточные мосты необходимы для фертильности [40].Они обеспечивают непрерывность цитоплазмы и достаточно велики (диаметр в диапазоне 0,2–10 мкм), чтобы позволить органеллам и/или макромолекулам проходить сквозь них [41]. Актин и анилин являются общими компонентами IBs, в то время как тубулин был обнаружен только в IBs мужской зародышевой линии (рис. 2C) [40]. У многих насекомых IBs между женскими зародышевыми клетками обеспечивают направленный транспорт питательных веществ, чтобы способствовать росту одной из клеток, которая разовьется в ооцит, тогда как другие клетки, после внесения своего цитоплазматического содержимого в ооцит, втягиваются и умирают. 42].Такие IBs у Drosophila melanogaster называются кольцевыми каналами и растут от диаметра около 0,5–1,5 мкм в гермарии до примерно 10 мкм на поздних стадиях оогенеза [42][43]. По мере увеличения диаметра кольцевого канала изменяется и его длина (от 0,22 мкм на ранних стадиях до 1,87 мкм на поздних стадиях) [43], однако она остается достаточно короткой по сравнению с длиной ТНТ (5-200 мкм) ( Таблица 1) . Половые клетки самок млекопитающих также соединены ТК, однако их диаметр не превышает 1 мкм [40].IBs мужской зародышевой линии разных видов имеют одинаковый средний диаметр 1–1,5 мкм и способствуют коммуникации зародышевых клеток и совместному использованию цитоплазматических компонентов, тем самым синхронизируя митотические деления клеток и вступление в мейоз [40].
Актин и анилин являются общими компонентами IBs, в то время как тубулин был обнаружен только в IBs мужской зародышевой линии (рис. 2C) [40]. У многих насекомых IBs между женскими зародышевыми клетками обеспечивают направленный транспорт питательных веществ, чтобы способствовать росту одной из клеток, которая разовьется в ооцит, тогда как другие клетки, после внесения своего цитоплазматического содержимого в ооцит, втягиваются и умирают. 42].Такие IBs у Drosophila melanogaster называются кольцевыми каналами и растут от диаметра около 0,5–1,5 мкм в гермарии до примерно 10 мкм на поздних стадиях оогенеза [42][43]. По мере увеличения диаметра кольцевого канала изменяется и его длина (от 0,22 мкм на ранних стадиях до 1,87 мкм на поздних стадиях) [43], однако она остается достаточно короткой по сравнению с длиной ТНТ (5-200 мкм) ( Таблица 1) . Половые клетки самок млекопитающих также соединены ТК, однако их диаметр не превышает 1 мкм [40].IBs мужской зародышевой линии разных видов имеют одинаковый средний диаметр 1–1,5 мкм и способствуют коммуникации зародышевых клеток и совместному использованию цитоплазматических компонентов, тем самым синхронизируя митотические деления клеток и вступление в мейоз [40]. Интересно, что в дополнение к мостикам зародышевой линии у некоторых развивающихся беспозвоночных были описаны соматически стабильные IB. Морфология соматических IBs Drosophila отличается от таковой кольцевых каналов. Они имеют стабильный диаметр примерно от 0,25 до 1 мкм (что ближе к диаметру тротила 0.2 – 0,7 мкм), а длиной 0,40 мкм (что очень мало по сравнению с длиной тротила 5-200 мкм) (табл. 1) [44]. Считается, что соматические ИБ способствуют обмену цитоплазмой и, возможно, органеллами, облегчая тем самым межклеточную коммуникацию, синхронизацию клеточного деления или дифференцировки и координацию клеточного поведения в процессе развития [40].
Интересно, что в дополнение к мостикам зародышевой линии у некоторых развивающихся беспозвоночных были описаны соматически стабильные IB. Морфология соматических IBs Drosophila отличается от таковой кольцевых каналов. Они имеют стабильный диаметр примерно от 0,25 до 1 мкм (что ближе к диаметру тротила 0.2 – 0,7 мкм), а длиной 0,40 мкм (что очень мало по сравнению с длиной тротила 5-200 мкм) (табл. 1) [44]. Считается, что соматические ИБ способствуют обмену цитоплазмой и, возможно, органеллами, облегчая тем самым межклеточную коммуникацию, синхронизацию клеточного деления или дифференцировки и координацию клеточного поведения в процессе развития [40].
–
РИСУНОК 2: Сходства и различия между ТНТ, цитонемами и межклеточными мостиками на уровне оптической микроскопии. ТНТ (А) и цитонемы (Б) имеют сходную морфологию, они содержат актин, но не тубулин. Однако TNT имеют открытые концы и обеспечивают непрерывность цитоплазмы, в то время как цитонемы имеют закрытые концы и допускают белок-белковые взаимодействия. |
 Межклеточные мосты (C) имеют открытые концы, но обычно короче и шире, чем TNT. Сообщалось также, что некоторые межклеточные мостики содержат тубулин. Все три типа клеточных структур способны переносить грузы. Однако если цитонемы переносят на своей поверхности везикулы, то ТНТ и межклеточные мостики способны переносить в свой просвет такие органеллы, как митохондрии.Поскольку диаметр ТНТ обычно меньше диаметра митохондрий, для переноса требуется выпячивание мембраны.
Межклеточные мосты (C) имеют открытые концы, но обычно короче и шире, чем TNT. Сообщалось также, что некоторые межклеточные мостики содержат тубулин. Все три типа клеточных структур способны переносить грузы. Однако если цитонемы переносят на своей поверхности везикулы, то ТНТ и межклеточные мостики способны переносить в свой просвет такие органеллы, как митохондрии.Поскольку диаметр ТНТ обычно меньше диаметра митохондрий, для переноса требуется выпячивание мембраны.–
В большинстве случаев отличная морфология IB облегчает их отличие от TNT (рис. 2) . Межклеточные мосты обычно имеют более короткие и толстые размеры, соединяя соседние клетки, в то время как TNT обычно тонкие и длинные и, таким образом, могут соединять клетки на больших расстояниях. Тем не менее, недавнее исследование развития гаструлы рыбок данио показало, что IB также способны соединять клетки на больших расстояниях (см. ниже) [45]. В этом свете различие в механизмах образования становится более важным критерием для различения этих двух структур. В то время как межклеточные мостики образуются между делящимися клетками в результате неполного цитокинеза, ТНТ не являются результатом незавершенного цитокинеза и образуются de novo между двумя отдельными клетками либо путем роста филоподиоподобного выпячивания [1], либо путем смещения двух прикрепленных клеток. оставив тротил в качестве троса [17] (рис. 3) . Таким образом, хотя предлагаемые функции IB могут пересекаться с предполагаемыми функциями TNT во время разработки, они явно представляют разные структуры, и, следовательно, их назначение также должно быть другим.В частности, TNT также могут соединять клетки различного происхождения. Это может происходить, например, в случае филоподиальных связей, наблюдаемых в процессе развития между эктодермальными и мезенхимальными клетками в гаструле морского ежа [46] или между клетками внутренней клеточной массы и клетками пристеночной трофэктодермы в бластоцисте мыши [47].
В этом свете различие в механизмах образования становится более важным критерием для различения этих двух структур. В то время как межклеточные мостики образуются между делящимися клетками в результате неполного цитокинеза, ТНТ не являются результатом незавершенного цитокинеза и образуются de novo между двумя отдельными клетками либо путем роста филоподиоподобного выпячивания [1], либо путем смещения двух прикрепленных клеток. оставив тротил в качестве троса [17] (рис. 3) . Таким образом, хотя предлагаемые функции IB могут пересекаться с предполагаемыми функциями TNT во время разработки, они явно представляют разные структуры, и, следовательно, их назначение также должно быть другим.В частности, TNT также могут соединять клетки различного происхождения. Это может происходить, например, в случае филоподиальных связей, наблюдаемых в процессе развития между эктодермальными и мезенхимальными клетками в гаструле морского ежа [46] или между клетками внутренней клеточной массы и клетками пристеночной трофэктодермы в бластоцисте мыши [47]. Потребуются дополнительные исследования, чтобы выяснить, являются ли филоподиальные связи, установленные в этих случаях между различными типами клеток, настоящими ТНТ (см. ниже).
Потребуются дополнительные исследования, чтобы выяснить, являются ли филоподиальные связи, установленные в этих случаях между различными типами клеток, настоящими ТНТ (см. ниже).
–
РИСУНОК 3: Механизмы образования ТНТ, цитонем и межклеточных мостиков по данным оптической живой микроскопии. ТНТ могут образовываться путем смещения клеток (A) , когда клетки сначала вступают в контакт, а затем образуют нанотрубку, мигрируя в противоположных направлениях. Морфологически это напоминает образование межклеточных мостиков (Б) . Однако межклеточные мостики образуются только между делящимися клетками путем незавершенного цитокинеза, тогда как ТНТ могут образовываться клетками различного происхождения. Таким образом, образование TNT требует слияния мембран до смещения клеток, тогда как образование межклеточных мостиков требует ингибирования абсциссии после клеточного деления.Другой механизм образования TNT, называемый «управляемым актином», имеет место, когда одна (C) или обе (E) клетки индуцируют рост филоподий-подобных выпячиваний. |
 Аналогичный механизм показан для цитонем. Цитонемы могут образовываться от продуцирующих к принимающим клеткам и наоборот (D), а также из них обоих (F). Однако в случае цитонемы разрастание завершается установлением контакта, а для образования ТНТ требуется последующее слияние мембран.
Аналогичный механизм показан для цитонем. Цитонемы могут образовываться от продуцирующих к принимающим клеткам и наоборот (D), а также из них обоих (F). Однако в случае цитонемы разрастание завершается установлением контакта, а для образования ТНТ требуется последующее слияние мембран.–
Цитонемы представляют собой другой тип клеточных выпячиваний, морфологически близких к ТНТ, но обладающих отличной функцией (Таблица 1) . Следовательно, их можно спутать с ТНТ, если отсутствует функциональная характеристика. В следующих разделах мы подробно обсудим их роль и сходство с TNT.
–
ТАБЛИЦА 1. Основные характеристики туннелирующих нанотрубок, межклеточных мостиков и цитонем. |
впервые были отмечены как длинные клеточные отростки, которые выступают из клеток имагинального диска крыльев дрозофилы . Эти выпячивания содержали актин, но не микротрубочки, имели длину в несколько диаметров клетки (до 700 мкм×03BC;м) и максимальный диаметр 200 нм. Их назвали цитонемами («клеточными нитями»), чтобы выделить их как филоподии особого типа [48]. Но определяющей характеристикой этих выпячиваний было то, что, независимо от расположения клеток в зачатке крыла, они были однородно ориентированы по направлению к срединной линии диска, где экспрессируется сигнальный белок морфогена Decapentaplegic (Dpp).Присутствие таких длинных филоподий, которые простираются между клетками, принимающими морфоген крылового диска, и клетками, экспрессирующими морфоген, свидетельствует об альтернативной возможности основанных на диффузии моделей дисперсии морфогена [48][49].
Эти выпячивания содержали актин, но не микротрубочки, имели длину в несколько диаметров клетки (до 700 мкм×03BC;м) и максимальный диаметр 200 нм. Их назвали цитонемами («клеточными нитями»), чтобы выделить их как филоподии особого типа [48]. Но определяющей характеристикой этих выпячиваний было то, что, независимо от расположения клеток в зачатке крыла, они были однородно ориентированы по направлению к срединной линии диска, где экспрессируется сигнальный белок морфогена Decapentaplegic (Dpp).Присутствие таких длинных филоподий, которые простираются между клетками, принимающими морфоген крылового диска, и клетками, экспрессирующими морфоген, свидетельствует об альтернативной возможности основанных на диффузии моделей дисперсии морфогена [48][49].
–
Морфогены определяются как формообразующие вещества, которые служат для генерации позиционной информации у эмбриона. Согласно модели, предложенной Lewis Wolpert, морфогены образуют градиенты концентрации внутри ткани, и специфический порог каждого морфогена определяет клеточную идентичность [50]. Классическая точка зрения состоит в том, что градиент концентрации образуется за счет простой диффузии. Однако модель диффузии, по-видимому, плохо применима для различных морфогенов, как в случае Wnt [подробно обсуждается в 51]. Фактически, лиганды Wnt посттрансляционно липидируются, что приводит к образованию плохо растворимой гидрофобной молекулы. Численное моделирование предполагает, что механизмам диффузии требуется несколько десятков часов, чтобы установить стабильный градиент морфогена и обеспечить соответствующий тканевый паттерн.Напротив, морфогенетический градиент Wnt действует в течение нескольких часов гаструляции. Более того, гаструльная стадия развития эмбриона характеризуется высокоинтенсивными клеточными реорганизациями, которые, вероятно, препятствуют градиентам передачи сигналов, формируемым внеклеточной диффузией [51]. В этом контексте цитонемы, по-видимому, являются вероятным механизмом для установления градиентов морфогена не только у Drosophila, но также и у позвоночных.
Классическая точка зрения состоит в том, что градиент концентрации образуется за счет простой диффузии. Однако модель диффузии, по-видимому, плохо применима для различных морфогенов, как в случае Wnt [подробно обсуждается в 51]. Фактически, лиганды Wnt посттрансляционно липидируются, что приводит к образованию плохо растворимой гидрофобной молекулы. Численное моделирование предполагает, что механизмам диффузии требуется несколько десятков часов, чтобы установить стабильный градиент морфогена и обеспечить соответствующий тканевый паттерн.Напротив, морфогенетический градиент Wnt действует в течение нескольких часов гаструляции. Более того, гаструльная стадия развития эмбриона характеризуется высокоинтенсивными клеточными реорганизациями, которые, вероятно, препятствуют градиентам передачи сигналов, формируемым внеклеточной диффузией [51]. В этом контексте цитонемы, по-видимому, являются вероятным механизмом для установления градиентов морфогена не только у Drosophila, но также и у позвоночных. В самом деле, было показано, что цитонемы Wnt эмбрионов рыбок данио обеспечивают быструю активацию передачи сигналов и формирование градиента в клетке-реципиенте в течение нескольких минут [52].По сравнению с формированием градиента на основе диффузии, в случае цитонем, градиенты Wnt могут зависеть от длины цитонемы и от частоты контакта с клетками-реципиентами. Клетки, расположенные ближе к источнику лиганда, чаще контактируют с цитонемами Wnt, чем клетки, расположенные дальше [51].
В самом деле, было показано, что цитонемы Wnt эмбрионов рыбок данио обеспечивают быструю активацию передачи сигналов и формирование градиента в клетке-реципиенте в течение нескольких минут [52].По сравнению с формированием градиента на основе диффузии, в случае цитонем, градиенты Wnt могут зависеть от длины цитонемы и от частоты контакта с клетками-реципиентами. Клетки, расположенные ближе к источнику лиганда, чаще контактируют с цитонемами Wnt, чем клетки, расположенные дальше [51].
–
Было показано, что в дополнение к морфогенам Dpp и Wnt цитонемы вносят вклад в передачу сигнала в факторах роста фибробластов (FGF), эпидермальном факторе роста (EGF), Wingless (Wg), Hedgehog (Hh) и путях Delta/Notch у дрозофилы [обзор в [29][49][51]].Более того, в дополнение к переносу Wnt у эмбрионов рыбок данио [52][53] сообщалось о переносе на основе цитонемы Shh (sonic hedgehog) и BMP7 у куриных эмбрионов [54][55]. Интересно, что было показано, что клетки могут расширять цитонемы, специфичные к разным морфогенам, и, таким образом, могут реагировать на множественные сигналы. В частности, было показано, что клетки зачатка воздушного мешка трахеи (ASP) дрозофилы расширяют цитонемы, содержащие Dpp-рецептор Tkv, в сторону близлежащих Dpp-продуцирующих клеток крылового диска, а также направляют FGFR-содержащие цитонемы в сторону клеток крылового диска, которые экспрессируют FGF.Примечательно, что исследованные цитонемы содержали только один тип рецептора, либо Tkv, либо FGFR, что означает, что каждая цитонема специфична только для одного лиганда [56]. Таким образом, цитонемы могут представлять собой высокоспецифичное средство прямой межклеточной коммуникации в процессе развития.
В частности, было показано, что клетки зачатка воздушного мешка трахеи (ASP) дрозофилы расширяют цитонемы, содержащие Dpp-рецептор Tkv, в сторону близлежащих Dpp-продуцирующих клеток крылового диска, а также направляют FGFR-содержащие цитонемы в сторону клеток крылового диска, которые экспрессируют FGF.Примечательно, что исследованные цитонемы содержали только один тип рецептора, либо Tkv, либо FGFR, что означает, что каждая цитонема специфична только для одного лиганда [56]. Таким образом, цитонемы могут представлять собой высокоспецифичное средство прямой межклеточной коммуникации в процессе развития.
–
В связи с морфологическим сходством ТНТ и цитонем, а также предположительно высокой значимостью цитонем в процессе развития мы считаем необходимым четкое отграничение ТНТ от цитонем при изучении функций ТНТ в процессе развития.Чтобы лучше понять сходства и различия между цитонемами и ТНТ, мы подробно обсудим эту тему в следующем разделе.
ТНТ И ЦИТОНЕМЫ: СХОДСТВА И РАЗЛИЧИЯ Как описано ранее, и TNT, и цитонемы представляют собой длинные и тонкие выросты на основе F-актина, поэтому морфологически они выглядят очень похожими. Наиболее заметной особенностью, отличающей эти два типа выпячивания клеток, является то, что цитонемы имеют закрытые концы, тогда как TNT являются открытыми (Рис. 2) .Следовательно, еще одно ключевое различие заключается в их функции: в то время как цитонемы обеспечивают передачу сигнала посредством белок-белковых взаимодействий [49], TNT позволяют передавать грузы посредством установления непрерывности цитоплазмы [1][57].
Наиболее заметной особенностью, отличающей эти два типа выпячивания клеток, является то, что цитонемы имеют закрытые концы, тогда как TNT являются открытыми (Рис. 2) .Следовательно, еще одно ключевое различие заключается в их функции: в то время как цитонемы обеспечивают передачу сигнала посредством белок-белковых взаимодействий [49], TNT позволяют передавать грузы посредством установления непрерывности цитоплазмы [1][57].
–
Учитывая, что функция переноса является ключевой особенностью ТНТ, необходимо подчеркнуть, что цитонемы также, по-видимому, переносят белки, хотя этот перенос, по-видимому, зависит от рецептора [28]. Интересно, что в пионерском исследовании цитонем дрозофилы авторы сообщили о перемещении везикул вдоль их поверхности (рис. 2В) [48].Совсем недавно было показано, что молекулы Shh переносятся в везикулах по поверхности цитонем дрозофилы [58]. Показано также, что цитонемы рыбок данио переносят Shh в частицах, ассоциированных с наружным листком мембраны цитонемы [54]. Эти наблюдения указывают на то, что морфология и способность переносить белки и/или везикулы могут быть недостаточными характеристиками, чтобы классифицировать выпячивание клетки как туннелирующую нанотрубку, хотя в случае ТНТ было продемонстрировано, что груз перемещается внутри трубки, а не вдоль мембраны. поверхность (рис. 2) [31].
Эти наблюдения указывают на то, что морфология и способность переносить белки и/или везикулы могут быть недостаточными характеристиками, чтобы классифицировать выпячивание клетки как туннелирующую нанотрубку, хотя в случае ТНТ было продемонстрировано, что груз перемещается внутри трубки, а не вдоль мембраны. поверхность (рис. 2) [31].
–
Поскольку и цитонемы, и TNT представляют собой выпячивания на основе F-актина, кажется вероятным рекрутирование одних и тех же актиновых регуляторов. Действительно, показано, что Myosin X ассоциирован как с цитонемами [52], так и с ТНТ [38]. Однако в случае пути, индуцирующего филоподии CDC42/IRSp53 [59][60], было показано, что хотя эти молекулы способны индуцировать образование цитонемы [52], они, по-видимому, ингибируют образование ТНТ [30]. Тем не менее, следует отметить, что было показано, что CDC42 индуцирует ТНТ в ненейрональных клеточных линиях [61].Это говорит о том, что механизмы образования ТНТ могут различаться в разных клетках, поэтому картина оказывается более сложной, чем предполагалось.
–
Как для цитонем, так и для ТНТ клетки-доноры и реципиенты способны способствовать их образованию (рис. 3) . Показано, что цитонемы исходят из клеток-продуцентов в клетки-получатели [52], из клеток-получателей в клетки-продуценты [48], а также из них обоих, что приводит к двукратному удлинению сигнального моста [51] ( Рисунок 3D , Ф) .Элегантный механизм образования и функционирования цитонем недавно был предложен для Wnt8a-цитонем у рыбок данио [53]. Было показано, что Wnt8a активирует как путь PCP (плоская клеточная полярность) путем взаимодействия с Ror2, так и путь β-катенина путем взаимодействия с Lrp6. Это позволяет Wnt8a регулировать образование цитонем и собственное размножение. В исходных клетках Wnt8a связывается и активирует путь PCP. Wnt/PCP влияет на движение конвергентного расширения и активирует GTPase CDC42, что приводит к разрастанию сигнальных филоподий.Затем Wnt8a загружается в эти цитонемы и транспортируется через ткань для активации пути β-катенина в реагирующих клетках, что приводит к индукции гена-мишени [53].
–
С другой стороны, ТНТ могут быть образованы двумя различными механизмами. Первый, названный актин-управляемым механизмом выпячивания, основан на способности одной или обеих клеток, участвующих в межклеточном контакте, индуцировать рост филоподий-подобных выпячиваний ( Figure 3C , E ).Затем за удлинением должно следовать установление контакта и слияние мембран [1][3]. Второй механизм образования ТНТ основан на смещении клеток (рис. 3А) , когда клетки сначала вступают в контакт, а затем образуют нанотрубку, мигрируя в противоположных направлениях [3][17]. Было описано несколько путей, включающих ремоделирование актина, Ras-GTPases, M-Sec, Rab-белки, а также p53 и EGFR, которые способствуют образованию TNT в различных клеточных контекстах, предполагая также специфичность типа клеток [30][38][62][63]. [64].Эти механизмы были подробно описаны в другом месте [обзор в [5][61][65]]. Более того, недавнее исследование, проведенное в нашей лаборатории, показало, что образование ТНТ в развивающихся нейронах и нервных клетках может стимулироваться путем Wnt. Однако, в отличие от цитонем, TNT образуются после активации Wnt ниже Ca 2+ кальмодулина, но не после β-катенинового пути [53][66]. Однако на сегодняшний день молекулярные детали механизма образования TNT в значительной степени неясны, особенно молекулярный механизм, опосредующий слияние клеток с клетками, все еще неясен.
Однако, в отличие от цитонем, TNT образуются после активации Wnt ниже Ca 2+ кальмодулина, но не после β-катенинового пути [53][66]. Однако на сегодняшний день молекулярные детали механизма образования TNT в значительной степени неясны, особенно молекулярный механизм, опосредующий слияние клеток с клетками, все еще неясен.
–
Интересно, что многочисленные стимулы, вызывающие образование ТНТ, могут быть определены как «стресс». Широкий спектр стрессов, включая H 2 O 2 , УФ, воспалительные состояния, вирусную инфекцию, агрегацию прионов, голодание в сыворотке и высокую концентрацию глюкозы, может вызвать образование ТНТ [61, 63, 67, 68]. [69]. Этот факт предполагает, что образование ТНТ может представлять собой тип реакции на стресс. Как только поврежденные клетки соединяются со здоровыми клетками, цитоплазматический перенос через TNT может восстановить их фенотип и продлить выживание [28].С другой стороны, ТНТ и ТНТ-подобные структуры также часто наблюдаются в стволовых, раковых и развивающихся клетках [4][70][71]. Это может указывать на то, что недифференцированный статус также может способствовать образованию ТНТ. Это указывает на еще одну возможную функцию ТНТ – как средства детерминации клеточных судеб. В целом эти наблюдения подтверждают гипотезу о ТНТ как о типе межклеточной коммуникации: будь то внешний сигнал, стресс или процесс развития, все это условия изменяющейся среды, условия, когда общение действительно необходимо и когда ТНТ легко образуются.Потребуются дополнительные исследования, чтобы опровергнуть эту гипотезу и точно определить молекулярные сигналы и компоненты, участвующие в образовании ТНТ как в физиологических, так и в патологических условиях.
Это может указывать на то, что недифференцированный статус также может способствовать образованию ТНТ. Это указывает на еще одну возможную функцию ТНТ – как средства детерминации клеточных судеб. В целом эти наблюдения подтверждают гипотезу о ТНТ как о типе межклеточной коммуникации: будь то внешний сигнал, стресс или процесс развития, все это условия изменяющейся среды, условия, когда общение действительно необходимо и когда ТНТ легко образуются.Потребуются дополнительные исследования, чтобы опровергнуть эту гипотезу и точно определить молекулярные сигналы и компоненты, участвующие в образовании ТНТ как в физиологических, так и в патологических условиях.
Поскольку ТНТ считаются средством межклеточной коммуникации, можно было бы ожидать, что они образуются, когда такая коммуникация необходима, а именно в меняющейся среде и в многоклеточном организме.В этом контексте модели развития представляют собой уникальную возможность исследовать нативные ТНТ в физиологических условиях.
–
Из обсуждаемой выше литературы становится ясно, что ТНТ, цитонемы и ИБ представляют собой разные структуры [33][40]. Однако во многих случаях оказывается затруднительным однозначно категоризировать выявленное выпячивание. Аналогичным образом, в нескольких исследованиях сообщалось о наличии длинных тонких выступов во время эмбрионального развития у разных видов позвоночных (таблица 2) [45, 46, 47, 72, 73, 74, 75].Интересно, что выпячивания, наблюдаемые в этих исследованиях, в дальнейшем называли «ТНТ-подобными» [4] или «цитонемоподобными» [29]. Однако ни одно из этих исследований не подтвердило наличие в их моделях функциональных ТНТ или цитонем. Некоторые из них скорее предположили наличие ПП необычной морфологии. Здесь мы обсуждаем исследования, описывающие структуры, которые не были четко классифицированы, и сравниваем их с «каноническими» ТНТ.
–
–
Первое описание ТНТ/цитонемоподобных структур у эмбриона было сделано в 1995 г. , когда Миллер и его коллеги сообщили о наличии тонких актинсодержащих филоподий, отходящих от эктодермальных клеток, клеток первичной мезенхимы и клеток вторичной мезенхимы в гаструле морского ежа [46]. ].Тонкие филоподии, расширенные клетками эктодермы, были короткими (5-10 мкм) и недолговечными по сравнению с расширенными клетками мезенхимы. Клетки первичной мезенхимы (ПМК) образовывали филоподии средней длины от 12 до 20 мкм, при этом отчетливые филоподии достигали длины более 80 мкм. Диаметр составлял примерно 200-400 нм, хотя однородность диаметра тонких филоподий иногда прерывалась утолщением, направляющимся к телу клетки. Показано, что в миграционную фазу ПМК вытягивают несколько тонких филоподий, тогда как наибольшее обилие тонких филоподий появляется после окончания активной миграции ПМК.Более того, тонкие филоподии преобладали в положениях эмбриона и в то время, когда более ранние исследования показали, что PMC взаимодействуют либо с эктодермой, либо с клетками вторичной мезенхимы для передачи информации о паттерне или клонах.
, когда Миллер и его коллеги сообщили о наличии тонких актинсодержащих филоподий, отходящих от эктодермальных клеток, клеток первичной мезенхимы и клеток вторичной мезенхимы в гаструле морского ежа [46]. ].Тонкие филоподии, расширенные клетками эктодермы, были короткими (5-10 мкм) и недолговечными по сравнению с расширенными клетками мезенхимы. Клетки первичной мезенхимы (ПМК) образовывали филоподии средней длины от 12 до 20 мкм, при этом отчетливые филоподии достигали длины более 80 мкм. Диаметр составлял примерно 200-400 нм, хотя однородность диаметра тонких филоподий иногда прерывалась утолщением, направляющимся к телу клетки. Показано, что в миграционную фазу ПМК вытягивают несколько тонких филоподий, тогда как наибольшее обилие тонких филоподий появляется после окончания активной миграции ПМК.Более того, тонкие филоподии преобладали в положениях эмбриона и в то время, когда более ранние исследования показали, что PMC взаимодействуют либо с эктодермой, либо с клетками вторичной мезенхимы для передачи информации о паттерне или клонах. Учитывая этот факт и то, что образование наблюдаемых филоподий не было связано с миграцией, авт. предположили сигнальную функцию этих выпячиваний. Более того, авторы предположили, что, учитывая тонкую и длинную морфологию филоподий, диффузия будет слишком медленной для передачи сигнала, и, таким образом, более вероятно, что будет задействован активный процесс, такой как электрические стимулы или каскад вторичных мессенджеров.Авторы также предположили, что ретроградный транспорт может обеспечивать активные средства клеточной коммуникации, о чем свидетельствуют выпячивания, которые наблюдались при движении от кончика филоподий к телу клетки [46]. Иными словами, авторы предполагали участие цитонем и/или ТНТ в клеточных коммуникациях у эмбрионов морского ежа еще до того, как были фактически обозначены термины «цитонемы» и «ТНТ».
Учитывая этот факт и то, что образование наблюдаемых филоподий не было связано с миграцией, авт. предположили сигнальную функцию этих выпячиваний. Более того, авторы предположили, что, учитывая тонкую и длинную морфологию филоподий, диффузия будет слишком медленной для передачи сигнала, и, таким образом, более вероятно, что будет задействован активный процесс, такой как электрические стимулы или каскад вторичных мессенджеров.Авторы также предположили, что ретроградный транспорт может обеспечивать активные средства клеточной коммуникации, о чем свидетельствуют выпячивания, которые наблюдались при движении от кончика филоподий к телу клетки [46]. Иными словами, авторы предполагали участие цитонем и/или ТНТ в клеточных коммуникациях у эмбрионов морского ежа еще до того, как были фактически обозначены термины «цитонемы» и «ТНТ».
–
Исследование Salas-Vidal and Lomeli 2004 представляет собой другое исследование, в котором предполагалось присутствие TNTs и/или цитонем во время развития без фактического использования этих терминов [47]. Эти авторы изучали возможные пути коммуникации между клетками внутренней клеточной массы (ICM) и клетками пристеночной трофэктодермы (mTE) в культуре целой мышиной бластоцисты. Во время роста бластоцисты пролиферация клеток в трофэктодерме (TE) зависит от взаимодействия с ICM и, таким образом, постепенно ограничивается полярной областью, которая остается близкой к ICM. Однако некоторые ТЕ-клетки в муральной области, потерявшие контакт с клетками ВКМ в ходе бластуляции, еще какое-то время делятся.Авторы задались вопросом, как эти удаленные клетки могут получать пролиферирующие сигналы от ICM. На самом деле им удалось обнаружить длинные клеточные выступы, проходящие через бластоцель и связывающие клетки ICM с клетками mTE. Выступы были прямыми и длинными (до 34,6 мкмx03BC;м), длина которых зависела от степени расширения бластоцеля. Эти перекрещивающиеся филоподии имели диаметр 200-400 нм в самом узком месте и содержали цитоплазму и актин. Авторы также смогли идентифицировать выпуклости, которые перемещались вдоль проекции и напоминали везикулы, которые демонстрировали ретроградное движение с отчетливой скачкообразной и непрерывной траекторией назад и вперед.
Эти авторы изучали возможные пути коммуникации между клетками внутренней клеточной массы (ICM) и клетками пристеночной трофэктодермы (mTE) в культуре целой мышиной бластоцисты. Во время роста бластоцисты пролиферация клеток в трофэктодерме (TE) зависит от взаимодействия с ICM и, таким образом, постепенно ограничивается полярной областью, которая остается близкой к ICM. Однако некоторые ТЕ-клетки в муральной области, потерявшие контакт с клетками ВКМ в ходе бластуляции, еще какое-то время делятся.Авторы задались вопросом, как эти удаленные клетки могут получать пролиферирующие сигналы от ICM. На самом деле им удалось обнаружить длинные клеточные выступы, проходящие через бластоцель и связывающие клетки ICM с клетками mTE. Выступы были прямыми и длинными (до 34,6 мкмx03BC;м), длина которых зависела от степени расширения бластоцеля. Эти перекрещивающиеся филоподии имели диаметр 200-400 нм в самом узком месте и содержали цитоплазму и актин. Авторы также смогли идентифицировать выпуклости, которые перемещались вдоль проекции и напоминали везикулы, которые демонстрировали ретроградное движение с отчетливой скачкообразной и непрерывной траекторией назад и вперед. Таким образом, эти выступы могут представлять ТНТ, впервые описанные Гердесом и его сотрудниками в том же году [1]. Однако авт. также показали сигнальные или цитонемные характеристики этих выпячиваний. Фактически, они обнаружили присутствие рецепторов EGF ErbB3 и рецепторов FGFR2 на выступах, которые исходят от TE-клеток. Присутствие этих рецепторов в проходящих филоподиях важно, учитывая, что их лиганды EGF и FGF4 способны стимулировать пролиферацию TE-клеток и что известно, что некоторые mTE-клетки все еще делятся, когда они находятся на расстоянии от ICM.Авторы предположили, что клетки ICM могут способствовать пролиферации клеток mTE посредством передачи сигналов EGF и FGF через длинные пересекающиеся филоподии [47]. В целом, это исследование предполагает существование выпячиваний, которые разделяют свойства цитонем и ТНТ, и поднимает вопрос, действительно ли цитонемы и ТНТ являются разными типами клеточных структур.
Таким образом, эти выступы могут представлять ТНТ, впервые описанные Гердесом и его сотрудниками в том же году [1]. Однако авт. также показали сигнальные или цитонемные характеристики этих выпячиваний. Фактически, они обнаружили присутствие рецепторов EGF ErbB3 и рецепторов FGFR2 на выступах, которые исходят от TE-клеток. Присутствие этих рецепторов в проходящих филоподиях важно, учитывая, что их лиганды EGF и FGF4 способны стимулировать пролиферацию TE-клеток и что известно, что некоторые mTE-клетки все еще делятся, когда они находятся на расстоянии от ICM.Авторы предположили, что клетки ICM могут способствовать пролиферации клеток mTE посредством передачи сигналов EGF и FGF через длинные пересекающиеся филоподии [47]. В целом, это исследование предполагает существование выпячиваний, которые разделяют свойства цитонем и ТНТ, и поднимает вопрос, действительно ли цитонемы и ТНТ являются разными типами клеточных структур.
–
Также в 2004 г. Teddy и Kulesa сообщили о наличии длинных и тонких выпячиваний у куриных эмбрионов при изучении миграции клеток нервного гребня в культурах целых эмбриональных эксплантатов [72]. Краниальный нервный гребень представляет собой высокоинвазивную субпопуляцию клеток. Тем не менее, клетки краниального нервного гребня не вторгаются, а скорее сортируются и мигрируют вдоль стереотипных потоков, формируя дальнейший паттерн периферических структур лица и шеи. Изучая клетки нервного гребня в потоках, авторы обнаружили две различные морфологии клеток и назвали их «биполярными клетками» и «волосатыми клетками». «Биполярные» клетки имели особенность иметь несколько длинных филоподий, которые обычно выстраивались в направлении вдоль траектории клетки.Эти филоподии были примерно 1 мкм в диаметре и имели длину 50-60 мкм, но могли достигать 100 мкм. Напротив, «волосатые» клетки имели большое количество филоподий, которые, по-видимому, не были распределены или выровнены в каком-либо определенном направлении и состояли как из длинных, так и из коротких отрезков, в диапазоне примерно от 20 мкм до 100 мкм. Интересно, что типичный поток клеток нервного гребня имел особенность «волосатых» клеток спереди и «биполярных» клеток в середине потока.
Краниальный нервный гребень представляет собой высокоинвазивную субпопуляцию клеток. Тем не менее, клетки краниального нервного гребня не вторгаются, а скорее сортируются и мигрируют вдоль стереотипных потоков, формируя дальнейший паттерн периферических структур лица и шеи. Изучая клетки нервного гребня в потоках, авторы обнаружили две различные морфологии клеток и назвали их «биполярными клетками» и «волосатыми клетками». «Биполярные» клетки имели особенность иметь несколько длинных филоподий, которые обычно выстраивались в направлении вдоль траектории клетки.Эти филоподии были примерно 1 мкм в диаметре и имели длину 50-60 мкм, но могли достигать 100 мкм. Напротив, «волосатые» клетки имели большое количество филоподий, которые, по-видимому, не были распределены или выровнены в каком-либо определенном направлении и состояли как из длинных, так и из коротких отрезков, в диапазоне примерно от 20 мкм до 100 мкм. Интересно, что типичный поток клеток нервного гребня имел особенность «волосатых» клеток спереди и «биполярных» клеток в середине потока. Было показано, что филоподии, образованные клетками нервного гребня в середине потока, устанавливают контакты между клетками.Этот контакт, по-видимому, приводил к тому, что следующая за ней клетка меняла направление своего движения в сторону нижестоящей клетки [72]. Эти данные свидетельствуют о том, что во время миграции клетки активно формируют филоподии не только для движения, но и для передачи сигналов. Учитывая тот факт, что клетки, как правило, начинали миграцию в потоке в «волосатой» морфологии, можно предположить, что при поиске исходной информации о направлении миграции клетки могли формировать многочисленные выпячивания в разные стороны.В дальнейшем клетки могут образовывать меньше выступов в направлении миграции, чтобы синхронизировать свою миграцию с другими клетками потока. Это исследование предполагает наличие сигнальных филоподий во время миграции клеток нервного гребня у куриных эмбрионов, которые могут представлять собой цитонемы или TNT. Однако авторы отмечают связь образования выпячиваний с делением клеток.
Было показано, что филоподии, образованные клетками нервного гребня в середине потока, устанавливают контакты между клетками.Этот контакт, по-видимому, приводил к тому, что следующая за ней клетка меняла направление своего движения в сторону нижестоящей клетки [72]. Эти данные свидетельствуют о том, что во время миграции клетки активно формируют филоподии не только для движения, но и для передачи сигналов. Учитывая тот факт, что клетки, как правило, начинали миграцию в потоке в «волосатой» морфологии, можно предположить, что при поиске исходной информации о направлении миграции клетки могли формировать многочисленные выпячивания в разные стороны.В дальнейшем клетки могут образовывать меньше выступов в направлении миграции, чтобы синхронизировать свою миграцию с другими клетками потока. Это исследование предполагает наличие сигнальных филоподий во время миграции клеток нервного гребня у куриных эмбрионов, которые могут представлять собой цитонемы или TNT. Однако авторы отмечают связь образования выпячиваний с делением клеток. Это может означать, что эти выпячивания скорее могут быть межклеточными мостиками, имеющими, однако, более протяженную морфологию, чем это обычно наблюдается.
Это может означать, что эти выпячивания скорее могут быть межклеточными мостиками, имеющими, однако, более протяженную морфологию, чем это обычно наблюдается.
–
В последующем исследовании той же группы [74] авторы изучали коммуникационные пути клеток нервного гребня у куриных эмбрионов, следя за движением фотоконвертированных флуоресцентных молекул. Авторы обнаружили, что в небольшом количестве из in vivo контактов клеток нервного гребня (около 13%) фотопреобразованные флуоресцентные молекулы перемещались из одной клетки нервного гребня в соседнюю клетку нервного гребня через тонкую (0,5–2 µ#x03BC;m широкий) сотовый мост. Напротив, в большинстве случаев (около 87%) фотопреобразованные флуоресцентные молекулы перемещались к средней точке клеточных расширений, определяя точку контакта между двумя соседями.Средняя длина клеточного моста составляла примерно 36,2 мкм x 03BC; м, а средняя ширина составляла 2,3 мкм x 03BC; м в средней точке между клетками. В большинстве наблюдений переноса цитоплазмы между клетками нервного гребня скорость переноса была выше, чем теоретическое значение случайной диффузии, что свидетельствует об активном механизме переноса [74]. Это исследование позволяет предположить, что как цитонемы (контакты без переноса цитоплазмы), так и ТНТ (контакты с переносом цитоплазмы) могли быть обнаружены в клетках нервного гребня куриного эмбриона в одни и те же сроки.Однако наблюдаемые выпячивания были толще обычных цитонем и ТНТ. Это может указывать либо на тот факт, что цитонемы и TNT могут иметь более изменчивую морфологию, чем предполагалось, либо на то, что наблюдаемые структуры представляют отдельный тип выпячиваний.
Это исследование позволяет предположить, что как цитонемы (контакты без переноса цитоплазмы), так и ТНТ (контакты с переносом цитоплазмы) могли быть обнаружены в клетках нервного гребня куриного эмбриона в одни и те же сроки.Однако наблюдаемые выпячивания были толще обычных цитонем и ТНТ. Это может указывать либо на тот факт, что цитонемы и TNT могут иметь более изменчивую морфологию, чем предполагалось, либо на то, что наблюдаемые структуры представляют отдельный тип выпячиваний.
–
В 2010 году Пиргаки и его коллеги использовали системы культивирования целых эмбрионов в сочетании с живой визуализацией генетически закодированного репортера для визуализации формирования нервной трубки у развивающегося эмбриона мыши, уделяя особое внимание последнему этапу процесса — слиянию нервных складок [73]. ].Авторы предположили, что они обнаружили в своей модели как цитонемоподобные, так и ТНТ-подобные структуры. Цитонемоподобные структуры представлены короткими и длинными (свыше 50 мкм x 03BC;m) гибкими клеточными отростками с бульбовидными концами, исходящими из ненейральных клеток эктодермы и располагающимися в промежутках между складками. ТНТ-подобные структуры были описаны как клеточные «мостики», перекрывающие промежуток между двумя нервными валиками в тот момент, когда они сближаются (≈20 µ#x03BC;m). Эти клеточные мостики были менее 1 мкм в ширину и содержали структуры ≈0.5 мкм в диаметре, которые были выделены миристоилированным репортером Венеры, что позволяет предположить везикулярную природу этих включений. Наблюдаемые мосты не считались IB, так как наблюдения предполагали их формирование de novo . В самом деле, было замечено, что эти структуры простираются над физическим промежутком до того, как складки сблизятся, и, следовательно, не могут представлять собой ни остатки клеточного деления, ни остатки, оставшиеся после разделения двух складок во время временного разделения [73].
ТНТ-подобные структуры были описаны как клеточные «мостики», перекрывающие промежуток между двумя нервными валиками в тот момент, когда они сближаются (≈20 µ#x03BC;m). Эти клеточные мостики были менее 1 мкм в ширину и содержали структуры ≈0.5 мкм в диаметре, которые были выделены миристоилированным репортером Венеры, что позволяет предположить везикулярную природу этих включений. Наблюдаемые мосты не считались IB, так как наблюдения предполагали их формирование de novo . В самом деле, было замечено, что эти структуры простираются над физическим промежутком до того, как складки сблизятся, и, следовательно, не могут представлять собой ни остатки клеточного деления, ни остатки, оставшиеся после разделения двух складок во время временного разделения [73].
–
В 2011 г. Caneparo и соавт. обнаружили присутствие IB-подобных структур, связывающих отдаленные пары клеток эпибласта во время ранних фаз гаструляции рыбок данио [45]. Характеристика этих межклеточных мостиков показала незначительные вариации по диаметру (до 1 мкмx03BC;м) и значительные вариации по длине, достигающие длины до 350 мкмx03BC;м при среднем пролете 215 мкмx03BC;м. Визуализация показала, что актин присутствовал по всей длине, тогда как тубулин был обнаружен только в проксимальной области IBs.Непрерывность мембраны была подтверждена фотопреобразованием связанного с мембраной белка Dendra в теле одной клетки и отслеживанием движений фотопреобразованного пула в направлении другой клетки. С помощью покадровой визуализации эмбрионов, экспрессирующих хроматиновый маркер Histone 2B, было показано, что IB сначала появляются между делящимися клетками на стадии бластулы, а затем сохраняются на протяжении гаструляции, морфогенетических движений и последующей клеточной митотической активности. Было обнаружено, что эти выпячивания довольно часто встречаются в гаструле; примерно одна из пяти клеток была наделена IBs.Такая высокая частота указывает на критические функции этих клеточных структур у эмбриона, однако было показано, что им не хватает какой-либо очевидной ориентации или предпочтительного положения на карте судеб, соединяя клетки как разных, так и одних и тех же эмбриональных областей [45].
Визуализация показала, что актин присутствовал по всей длине, тогда как тубулин был обнаружен только в проксимальной области IBs.Непрерывность мембраны была подтверждена фотопреобразованием связанного с мембраной белка Dendra в теле одной клетки и отслеживанием движений фотопреобразованного пула в направлении другой клетки. С помощью покадровой визуализации эмбрионов, экспрессирующих хроматиновый маркер Histone 2B, было показано, что IB сначала появляются между делящимися клетками на стадии бластулы, а затем сохраняются на протяжении гаструляции, морфогенетических движений и последующей клеточной митотической активности. Было обнаружено, что эти выпячивания довольно часто встречаются в гаструле; примерно одна из пяти клеток была наделена IBs.Такая высокая частота указывает на критические функции этих клеточных структур у эмбриона, однако было показано, что им не хватает какой-либо очевидной ориентации или предпочтительного положения на карте судеб, соединяя клетки как разных, так и одних и тех же эмбриональных областей [45]. Морфологические свойства этих выпячиваний вместе с непрерывностью мембраны напоминают признаки ТНТ. Отсутствие какой-либо явной направленности также не соответствовало ключевому признаку цитонем. Однако авторы также заметили, что образование этих выпячиваний было связано с клеточным делением, и, таким образом, пришли к выводу, что они могут представлять тип IB необычной морфологии, а не TNT.
Морфологические свойства этих выпячиваний вместе с непрерывностью мембраны напоминают признаки ТНТ. Отсутствие какой-либо явной направленности также не соответствовало ключевому признаку цитонем. Однако авторы также заметили, что образование этих выпячиваний было связано с клеточным делением, и, таким образом, пришли к выводу, что они могут представлять тип IB необычной морфологии, а не TNT.
–
И последнее, но не менее важное: в работе Danilchik et al. 2013 сообщалось о наличии исключительно длинных филоподий-подобных структур, охватывающих бластоцель ранней бластулы Xenopus laevis [75]. Эти структуры были организованы в виде массивов параллельных отростков, устойчиво соединяющих бластомеры на расстояниях до 250 мкм x 03BC;м. Эти соединения содержали актин, но не тубулин, имели диаметры в пределах от 200 до 700 нм и обеспечивали перенос мембранных везикул между клетками.Первоначально они образовались между сестринскими бластомерами и удлинились за счет разделения бластомеров, вызванного расширением бластоцеля. Эти проекции сохранялись в течение нескольких часов в течение нескольких циклов деления (т. е. от стадии 32 клеток до стадии 256 клеток). Хотя описанные выпячивания образовались между делящимися бластомерами, авторам удалось отличить их от IB благодаря их морфологии и специфическому расположению. На самом деле, авторы также описали бластомеры, которые все еще были соединены IB и в то же время проецировали многочисленные длинные филоподии в нескольких направлениях [75].Это наблюдение приводит нас к предположению, что разные типы выпячиваний могли образовываться одной и той же клеткой в одно и то же время, и не следует исключать возможности образования ТНТ и/или цитонемы при делении клеток. Следовательно, связь образования выпячиваний с делением клеток, показанная в описанных выше работах [45][72], не обязательно означает, что эти выпячивания не являются цитонемами или ТНТ.
Эти проекции сохранялись в течение нескольких часов в течение нескольких циклов деления (т. е. от стадии 32 клеток до стадии 256 клеток). Хотя описанные выпячивания образовались между делящимися бластомерами, авторам удалось отличить их от IB благодаря их морфологии и специфическому расположению. На самом деле, авторы также описали бластомеры, которые все еще были соединены IB и в то же время проецировали многочисленные длинные филоподии в нескольких направлениях [75].Это наблюдение приводит нас к предположению, что разные типы выпячиваний могли образовываться одной и той же клеткой в одно и то же время, и не следует исключать возможности образования ТНТ и/или цитонемы при делении клеток. Следовательно, связь образования выпячиваний с делением клеток, показанная в описанных выше работах [45][72], не обязательно означает, что эти выпячивания не являются цитонемами или ТНТ.
–
Все рассмотренные выше исследования показали существование цитонемоподобных и ТНТ-подобных структур in vivo во время развития эмбриона. Однако ни один из них не подтвердил функциональность и, следовательно, идентичность наблюдаемых структур. Необходим дальнейший функциональный анализ, чтобы подтвердить или опровергнуть присутствие ТНТ и/или цитонем в этих моделях и оценить возможную роль ТНТ во время развития.
Однако ни один из них не подтвердил функциональность и, следовательно, идентичность наблюдаемых структур. Необходим дальнейший функциональный анализ, чтобы подтвердить или опровергнуть присутствие ТНТ и/или цитонем в этих моделях и оценить возможную роль ТНТ во время развития.
Одна из целей этого обзора — вдохновить на дальнейшие исследования роли, а также функционального и структурного сходства ТНТ и других выступов в моделях развития.
–
Особый интерес представляет вопрос о том, существуют ли истинные ТНТ in vivo во время эмбрионального развития, поскольку, в отличие от цитонем, существование ТНТ в эмбрионе пока не показано. В то время как функции цитонем во время развития активно изучаются, возможные функции ТНТ во время развития на данный момент только предполагаются. Основное соображение заключается в том, что TNTs, электрически связывая клетки эмбриона, могут способствовать координации их миграционной активности и, таким образом, оркестровке морфогенных движений [4][5]. Однако в то же время, подобно межклеточным мостикам зародышевой линии, ТНТ могут способствовать синхронизации клеточных делений и запрограммированной клеточной гибели [40]. Действительно, было показано, что ТНТ переносят лиганд Fas сигнала смерти и активные каспазы in vitro и, таким образом, могут индуцировать апоптоз в принимающих клетках [13]. Другая возможность заключается в том, что TNT могут способствовать детерминации клеточных судеб. Это предположение подтверждается наблюдением in vitro , что TNT-подобные структуры, образованные между мезенхимальными стволовыми клетками и кардиомиоцитами или гладкомышечными клетками, необходимы для дифференцировки стволовых клеток в кардиомиоциты или гладкомышечные клетки, соответственно [76].Хотя такие функции ТНТ в развитии были предложены, фактические функции ТНТ, а также их существование в эмбрионе еще предстоит исследовать. Мы считаем, что новые методы живой визуализации и криоэлектронной микроскопии помогут ответить на эти вопросы, а также оценить морфологические и функциональные различия между ТНТ и другими клеточными выпячиваниями.
Однако в то же время, подобно межклеточным мостикам зародышевой линии, ТНТ могут способствовать синхронизации клеточных делений и запрограммированной клеточной гибели [40]. Действительно, было показано, что ТНТ переносят лиганд Fas сигнала смерти и активные каспазы in vitro и, таким образом, могут индуцировать апоптоз в принимающих клетках [13]. Другая возможность заключается в том, что TNT могут способствовать детерминации клеточных судеб. Это предположение подтверждается наблюдением in vitro , что TNT-подобные структуры, образованные между мезенхимальными стволовыми клетками и кардиомиоцитами или гладкомышечными клетками, необходимы для дифференцировки стволовых клеток в кардиомиоциты или гладкомышечные клетки, соответственно [76].Хотя такие функции ТНТ в развитии были предложены, фактические функции ТНТ, а также их существование в эмбрионе еще предстоит исследовать. Мы считаем, что новые методы живой визуализации и криоэлектронной микроскопии помогут ответить на эти вопросы, а также оценить морфологические и функциональные различия между ТНТ и другими клеточными выпячиваниями.
ССЫЛКИ
- Растом А., Саффрич Р., Маркович И., Вальтер П., Гердес Х.Х. ( 2004 ).Нанотрубчатые магистрали для межклеточного транспорта органелл. Наука 303(5660): 1007-1010. 10.1126/наука.1093133
- Бейкер М ( 2017 ). Линии связи. Природа 549(7672): 322-324. 10.1038/549322а
- Абунит С., Зурзоло С. ( 2012 ). Проводка через туннельные нанотрубки — от электрических сигналов до переноса органелл. J Cell Sci 125 (часть 5): 1089-1098. 10.1242/jcs.083279
- Гердес Х.Х., Растом А., Ван Х ( 2013 ).Туннельные нанотрубки, новый путь межклеточной коммуникации в разработке. Mech Dev 130(6-8): 381-387. 10.1016/j.mod.2012.11.006
- Сисахтнежад С., Хосрави Л. ( 2015 ). Возникающие физиологические и патологические последствия образования туннельных нанотрубок между клетками. Eur J Cell Biol 94(10): 429-443. 10.1016/j.ejcb.2015.06.010
- Виньяс М.Л., Кайседо А., Бронделло Дж.М., Йоргенсен С. ( 2017 ). Клеточные соединения путем туннелирования нанотрубок: влияние митохондриального трафика на метаболизм клеток-мишеней, гомеостаз и реакцию на терапию. Stem Cells Int 2017: 6917941. 10.1155/2017/6917941
- Хекматшоар Ю., Нахле Дж., Галлони М., Винье М.Л. ( 2018 ). Роль метаболизма и туннельного межклеточного обмена митохондрий, опосредованного нанотрубками, в лекарственной устойчивости рака. Biochem J 475(14): 2305-2328. 10.1042/bcj20170712
- Watkins SC, Salter RD ( 2005 ). Функциональная связь между иммунными клетками, опосредованная туннелирующими нанотрубочками. Иммунитет 23(3): 309-318.10.1016/j.иммуни.2005.08.009
- Тада М., Конча М.Л. ( 2001 ). Гаструляция позвоночных: кальциевые волны управляют движением клеток. Карр Биол 11(12): R470-472. 10.1016/s0960-9822(01)00284-6
- Левин М ( 2007 ). Связь щелевых соединений в морфогенезе. Prog Biophys Mol Biol 94(1-2): 186-206. 10.1016/j.pbiomolbio.2007.03.005
- Ван Х, Букорештлиев Н.В., Гердес Х.Х. ( 2012 ). Развивающиеся нейроны образуют временные нанотрубки, облегчающие электрическую связь и передачу сигналов кальция с отдаленными астроцитами. PLoS One 7(10): e47429. 10.1371/journal.pone.0047429
- Дюпон М., Сурьян С., Луго-Вилларино Г., Маридонно-Парини И., Веролле С. ( 2018 ). Туннельные нанотрубки: интимная связь между миелоидными клетками. Фронт Иммунол 9:43. 10.3389/fimmu.2018.00043
- Аркрайт П.Д., Лучетти Ф., Тур Дж., Робертс С., Аюб Р., Моралес А.П., Родригес Дж.Дж., Гилмор А., Канонико Б., Папа С., Эспости М.Д. ( 2010 ). Fas-стимуляция Т-лимфоцитов способствует быстрому межклеточному обмену сигналами смерти через мембранные нанотрубки. Cell Res 20(1): 72-88. 10.1038/кр.2009.112
- Лу Э., Фудзисава С., Морозов А., Барлас А., Ромин Ю., Доган Ю., Голами С., Морейра А.Л., Манова-Тодорова К., Мур М.А. ( 2012 ). Туннельные нанотрубки обеспечивают уникальный канал для межклеточного переноса клеточного содержимого при злокачественной мезотелиоме плевры человека. PLoS One 7(3): e33093. 10.1371/journal.pone.0033093
- Тайанити В., Диксон Э.Л., Стир С., Субраманиан С., Лу Э. ( 2014 ). Опухоль-стромальный перекрестный разговор: прямой перенос онкогенных микроРНК между клетками через туннельные нанотрубки. Перевод Res 164(5): 359-365. 10.1016/j.trsl.2014.05.011
- Освальд М., Юнг Э., Вик В., Винклер Ф. ( 2019 ). Туннельные структуры, подобные нанотрубкам, в опухолях головного мозга. Рак Rep 2019: e1181. 10.1002/номер 2.1181
- Совински С., Джолли С., Бернингхаузен О. , Пурбху М.А., Шово А., Колер К., Оддос С., Эйссманн П., Бродский Ф.М., Хопкинс С., Онфельт Б., Саттентау К., Дэвис Д.М. ( 2008 ). Мембранные нанотрубки физически соединяют Т-клетки на большие расстояния, представляя собой новый путь передачи ВИЧ-1. Nat Cell Biol 10(2): 211-219. 10.1038/ncb1682
- Хашимото М., Бхуян Ф., Хиёси М., Ноёри О., Нассер Х., Миядзаки М., Сайто Т., Кондо Ю., Осада Х., Кимура С., Хасэ К., Оно Х., Судзу С. ( 2016 ). Возможная роль образования туннельных нанотрубок в распространении ВИЧ-1 в макрофагах. J Immunol 196(4): 1832-1841. 10.4049/иммунол.1500845
- Гуссе К., Шифф Э., Ланжевен С., Мариянович З., Капуто А., Брауман Д.Т., Шенуар Н., де Шомон Ф., Мартино А., Эннинга Дж., Оливо-Марин Дж.С., Маннел Д., Зурзоло С. ( 2009 ).Прионы захватывают туннелирующие нанотрубки для межклеточного распространения. Nat Cell Biol 11(3): 328-336. 10.1038/ncb1841
- Абунит С. , Буссе Л., Лориа Ф., Чжу С., де Шомон Ф., Пьери Л., Оливо-Марин Дж. К., Мелки Р., Зурзоло С. ( 2016 ). Туннельные нанотрубки распространяют фибриллярный альфа-синуклеин посредством межклеточного переноса лизосом. Embo J 35(19): 2120-2138. 10.15252/прил.201593411
- Abounit S, Wu JW, Duff K, Victoria GS, Zurzolo C ( 2016 ).Туннельные нанотрубки: возможный путь распространения тау и других прионоподобных белков при нейродегенеративных заболеваниях. Прион 10(5): 344-351. 10.1080/19336896.2016.1223003
- Victoria GS, Zurzolo C ( 2017 ). Распространение прионоподобных белков лизосомами и туннельными нанотрубками: значение для нейродегенеративных заболеваний. J Cell Biol 216(9): 2633-2644. 10.1083/jcb.201701047
- Чжу С., Абунит С., Корт С., Зурзоло С. ( 2017 ).Перенос нарушенных при шизофрении 1 агрегатов между нейроноподобными клетками происходит в туннелирующих нанотрубках и стимулируется дофамином. Опен Биол 7(3): 160328. 10.1098/rsob.160328
- Дэвис Д.М., Совински С. ( 2008 ). Мембранные нанотрубки: динамические связи на большие расстояния между клетками животных. Nat Rev Mol Cell Biol 9(6): 431-436. 10.1038/nrm2399
- Хуртиг Дж., Чиу Д.Т., Онфельт Б. ( 2010 ). Межклеточные нанотрубки: выводы из исследований изображений и не только. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2(3): 260-276. 10.1002/wнан.80
- Austefjord MW, Gerdes HH, Wang X ( 2014 ). Туннельные нанотрубки: разнообразие морфологии и структуры. Коммун Интегр Биол 7(1): e27934. 10.4161/cib.27934
- Ариази Дж., Беновиц А., Де Биази В., Ден Бур М.Л., Черки С., Куй Х., Дуйе Н., Евгенин Э.А., Фавр Д., Гудман С., Гуссе К., Ханейн Д., Исраэль Д.И., Кимура С., Киркпатрик Р.Б., Кун Н., Чон С., Лу Э., Майярд Р., Майо С., Окафо Г., Освальд М., Паскье Дж., Полак Р. , Прадель Г., де Рой Б., Шеффер П., Скебердис В.А., Смит И.Ф., Танвир А. и др.( 2017 ). Туннельные нанотрубки и щелевые соединения — их роль в дальних межклеточных коммуникациях во время развития, здоровья и болезней. Front Mol Neurosci 10: 333. 10.3389/fnmol.2017.00333
- Ямашита Ю.М., Инаба М., Бущак М. ( 2018 ). Специализированные межклеточные коммуникации через цитонемы и нанотрубки. Annu Rev Cell Dev Biol 34: 59-84. 10.1146/annurev-cellbio-100617-062932
- Бущак М., Инаба М., Ямашита Ю.М. ( 2016 ).Сигнализация с помощью сотовых выступов: сохранение конфиденциальности разговора. Trends Cell Biol 26(7): 526-534. 10.1016/j.tcb.2016.03.003
- Делаж Э., Сервантес Д.С., Пенар Э., Шмитт С., Сян С., Дисанза А., Скита Г., Зурзоло С. ( 2016 ). Дифференциальная идентичность филоподий и туннельных нанотрубок проявляется в противоположных функциях актиновых регуляторных комплексов. Научный представитель 6: 39632. 10.1038/srep39632
- Сартори-Рупп А., Кордеро Сервантес Д., Пепе А., Делаж Э., Гуссе К., Корройер-Дульмон С., Шмитт С., Крейнсе-Локер Дж., Цурзоло С. ( 2019 ).Корреляционная криоэлектронная микроскопия выявляет структуру ТНТ в нейрональных клетках. Нацкоммун 10(1): 342. 10.1038/s41467-018-08178-7
- Абунит С., Делаж Э., Зурзоло С. ( 2015 ). Идентификация и характеристика туннелирующих нанотрубок для межклеточной торговли. Curr Protoc Cell Biol 67(12.10) 1-21. 10.1002/0471143030.cb1210s67
- Маттес Б., Шолпп С. ( 2018 ). Новая роль контактно-опосредованной клеточной коммуникации в развитии тканей и заболеваниях. Histochem Cell Biol 150(5): 431-442. 10.1007/s00418-018-1732-3
- Ван Х, Гердес Х.Х. ( 2015 ). Перенос митохондрий через туннелирующие нанотрубки спасает апоптотические клетки PC12. Cell Death Differ 22(7): 1181-1191. 10.1038/cdd.2014.211
- Онфельт Б., Недвецки С., Беннингер Р.К., Пурбху М.А., Совински С., Хьюм А.Н., Сибра М.С., Нил М.А., Френч П.М., Дэвис Д.М. ( 2006 ). Структурно различные мембранные нанотрубки между макрофагами человека поддерживают везикулярный трафик на большие расстояния или серфинг бактерий. J Иммунол 177(12): 8476-8483. 10.4049/иммунол.177.12.8476
- Ван X, Гердес HH ( 2012 ). Электрическая связь на большие расстояния через туннельные нанотрубки. Biochimica et biophysica acta 1818(8): 2082-2086. 10.1016/j.bbamem.2011.09.002.
- Бохил А.Б., Робертсон Б.В., Чейни Р.Е. ( 2006 ). Миозин-X представляет собой молекулярный мотор, участвующий в формировании филоподий. Proc Natl Acad Sci USA 103(33): 12411-12416. 10.1073/пнас.0602443103
- Гуссе К., Марзо Л., Коммер П.Х., Зурзоло С. ( 2013 ). Myo10 является ключевым регулятором образования ТНТ в нейрональных клетках. J Cell Sci 126 (Pt 19): 4424-4435. 10.1242/jcs.129239
- Гринбаум М.П., Ивамори Т., Бухольд Г.М., Мацук М.М. ( 2011 ). Межклеточные мостики зародышевых клеток. Колд Спринг Харб Перспект Биол 3(8): a005850. 10.1101/cshperspect.a005850
- Хаглунд К., Незис И.П., Стенмарк Х. ( 2011 ). Структура и функции стабильных межклеточных мостиков, образующихся в результате незавершенного цитокинеза в процессе развития. Коммун Интегр Биол 4(1): 1-9. 10.4161/см.4.1.13550
- Робинсон Д.Н., Кули Л. ( 1996 ). Стабильные межклеточные мосты в развитии: цитоскелет, выстилающий туннель. Trends Cell Biol 6(12): 474-479. 10.1016/0962-8924(96)84945-2.
- Робинсон Д.Н., Кули Л. ( 1997 ). Генетический анализ актинового цитоскелета яичника дрозофилы. Annu Rev Cell Dev Biol 13: 147-170. 10.1146/annurev.cellbio.13.1.147
- Тилни Л.Г., Тилни М.С., Гильдия Г.М. ( 1996 ).Формирование пучков актиновых филаментов в кольцевых каналах развивающихся фолликулов дрозофилы. J Cell Biol 133(1): 61-74. 10.1083/jcb.133.1.61
- Вудрафф Р.И., Тилни Л.Г. ( 1998 ). Межклеточные мосты между эпителиальными клетками в фолликулах яичников дрозофилы: возможная помощь в локализованной передаче сигналов. Dev Biol 200(1): 82-91. 10.1006/dbio.1998.8948
- Канепаро Л., Пантазис П., Демпси В., Фрейзер С.Е. ( 2011 ). Межклеточные мостики при гаструляции позвоночных. PLoS One 6(5): e20230. 10.1371/journal.pone.0020230
- Миллер Дж., Фрейзер С.Е., Макклей Д. ( 1995 ). Динамика тонких филоподий при гаструляции морского ежа. Разработка 121(8): 2501-2511. 7671814.
- Салас-Видаль Э., Ломели Х. ( 2004 ). Визуализация динамики филоподий в бластоцисте мыши. Dev Biol 265(1): 75-89. 10.1016/j.ydbio.2003.09.012
- Рамирес-Вебер Ф.А., Корнберг Т.Б. ( 1999 ). Цитонемы: клеточные отростки, проецирующиеся к основному сигнальному центру в имагинальных дисках дрозофилы. Сотовый 97(5): 599-607. 10.1016/s0092-8674(00)80771-0
- Корнберг Т.Б., Рой С. ( 2014 ). Цитонемы как специализированные сигнальные филоподии. Разработка 141(4): 729-736. 10.1242/dev.086223
- Вольперт Л. ( 1969 ). Позиционная информация и пространственный паттерн клеточной дифференцировки. J Theor Biol 25(1): 1-47. 10.1016/s0022-5193(69)80016-0
- Станганелло Э., Шолпп С. ( 2016 ). Роль цитонем в транспорте Wnt. J Cell Sci 129(4): 665-672. 10.1242/jcs.182469
- Станганелло Э., Хагеманн А.И., Маттес Б., Синнер С., Мейен Д. , Вебер С., Шуг А., Раз Э., Шолпп С. ( 2015 ). Транспорт Wnt на основе филоподий во время формирования паттерна ткани позвоночных. Нац Коммуна 6: 5846. 10.1038/ncomms6846
- Маттес Б., Данг Ю., Грейциус Г., Кауфманн Л.Т., Прунше Б., Розенбауэр Дж., Стегмайер Дж., Микут Р., Озбек С., Ниенхаус Г.У., Шуг А., Виршуп Д.М., Шолпп С. ( 2018 ). Wnt/PCP контролирует распространение сигналов Wnt/beta-catenin цитонемами у позвоночных. Элайф 7: e36953. 10.7554/eLife.36953
- Сандерс Т.А., Ллагостера Э., Барна М. ( 2013 ). Специализированные филоподии направляют транспорт SHH на большие расстояния во время формирования паттерна тканей позвоночных. Природа 497(7451): 628-632. 10.1038/природа12157
- Шлютер Дж., Микава Т. ( 2018 ). Развитие полостей тела управляется переносом морфогена между зародышевыми слоями. Cell Rep 24(6): 1456-1463. 10.1016/j.celrep.2018.07.015
- Рой С., Хсюн Ф., Корнберг Т.Б. ( 2011 ).Специфичность цитонем дрозофилы для различных сигнальных путей. Наука 332(6027): 354-358. 10.1126/наука.1198949
- Gerdes HH, Carvalho RN ( 2008 ). Межклеточный перенос, опосредованный туннелирующими нанотрубками. Curr Opin Cell Biol 20(4): 470-475. 10.1016/j.ceb.2008.03.005
- Градилья А.С., Гонсалес Э., Сейхо И., Андрес Г., Бишофф М., Гонсалес-Мендес Л., Санчес В., Кальехо А., Ибанес К., Герра М., Ортигао-Фариас Д.Р., Сазерленд Д.Д., Гонсалес М., Баррио Р., Фалькон-Перес Д.М. , Герреро I ( 2014 ).Экзосомы как переносчики Hedgehog в опосредованном цитонемами транспорте и секреции. Нац Коммуна 5: 5649. 10.1038/ncomms6649
- Disanza A, Mantoani S, Hertzog M, Gerboth S, Frittoli E, Steffen A, Berhoerster K, Kreienkamp HJ, Milanesi F, Fiore PPD, Ciliberto A, Stradal TEB, Scita G ( 2006 ). Регуляция формы клеток с помощью Cdc42 опосредуется синергической активностью связывания актина комплексом Eps8-IRSp53. Nat Cell Biol 8: 1337. 10.1038/ncb1502
- Дисанза А., Бизи С., Винтерхофф М., Миланези Ф., Ушаков Д.С., Каст Д., Маригетти П., Ромет-Лемонн Г., Мюллер Х.М., Никель В., Линкнер Дж., Уотершут Д., Ампе С., Кортеллино С., Паламидесси А., Домингес Р., Carlier MF, Faix J, Scita G ( 2013 ).CDC42 переключает IRSp53 с ингибирования роста актина на удлинение за счет кластеризации VASP. Embo J 32(20): 2735-2750. 10.1038/emboj.2013.208
- Кимура С., Хасэ К., Оно Х. ( 2013 ). Молекулярные основы индукции и формирования туннельных нанотрубок. Cell Tissue Res 352(1): 67-76. 10.1007/s00441-012-1518-1
- Хасэ К., Кимура С., Такацу Х., Омаэ М., Кавано С., Китамура Х., Ито М., Ватарай Х., Хазелетт К.С., Йеман С., Оно Х. ( 2009 ).M-Sec способствует образованию мембранных нанотрубок, взаимодействуя с Ral и экзоцистным комплексом. Nat Cell Biol 11(12): 1427-1432. 10.1038/ncb1990
- Ван И, Цуй Дж, Сун С, Чжан И ( 2011 ). Развитие туннельных нанотрубок в астроцитах зависит от активации p53. Cell Death Differ 18(4): 732-742. 10.1038/cdd.2010.147
- Чжу С., Бхат С., Сян С., Кучицу Ю., Фукуда М., Зурзоло С. ( 2018 ). Каскад Rab11a-Rab8a регулирует образование туннельных нанотрубок посредством рециркуляции везикул. J Cell Sci 131(19): jcs215889. 10.1242/jcs.215889
- Марзо Л., Гуссе К., Зурзоло С. ( 2012 ). Многогранная роль туннелирующих нанотрубок в межклеточной коммуникации. Фронт Физиол 3:72. 10.3389/Ффиз.2012.00072
- Варгас Дж.Ю., Лориа Ф., Ву Дж.В., Кордова Г., Нонака Т., Беллоу С., Сян С., Хасегава М., ван Вурден Г.М., Троллет С., Зурзоло С. ( 2019 ). Путь Wnt/Ca2+ участвует в межнейронной коммуникации, опосредованной туннелирующими нанотрубками. Embo J (2019) 38: e101230. 10.15252/прил.2018101230
- Zhu D, Tan KS, Zhang X, Sun AY, Sun GY, Lee JC ( 2005 ). Перекись водорода изменяет свойства мембран и цитоскелета, увеличивает межклеточные связи в астроцитах. J Cell Sci 118 (Pt 16): 3695-3703. 10.1242/jcs.02507
- Аггарвал А., Иемма Т.Л., Ши И., Ньюсом Т.П., Макаллери С., Каннингем А.Л., Тервилл С.Г. ( 2012 ). Мобилизация распространения ВИЧ через прозрачные 2-зависимые филоподии в инфицированных дендритных клетках. PLoS Pathog 8(6): e1002762. 10.1371/журнал.ppat.1002762
- Чжу С., Виктория Г.С., Марзо Л., Гош Р., Зурзоло С. ( 2015 ). Агрегаты прионов переходят через туннелирующие нанотрубки в эндоцитарные везикулы. Прион 9(2): 125-135. 10.1080/19336896.2015.1025189
- Лу Э., Голами С., Ромин Ю., Таянити В., Фудзисава С., Дезир С., Стир С.Дж., Субраманиан С., Фонг Ю. , Манова-Тодорова К., Мур MAS ( 2017 ). Визуализация туннельных мембранных трубок объясняет клеточную коммуникацию в опухолях. Trends Cancer 3(10): 678-685. 10.1016/j.trecan.2017.08.001
- Murray LMA, Краснодембская АД ( 2018 ). Краткий обзор: Межклеточная коммуникация посредством переноса органелл в биологии и терапевтических применениях стволовых клеток. Стволовые клетки 37(1): 14-25. 10.1002/шток.2922
- Тедди Дж. М., Кулеса П. М. ( 2004 ). In vivo доказательства ближней и дальней клеточной коммуникации в клетках краниального нервного гребня. Разработка 131(24): 6141-6151.10.1242/dev.01534
- Пиргаки С., Трейнор П., Хаджантонакис А.К., Нисвандер Л. ( 2010 ). Динамическая визуализация закрытия нервной трубки млекопитающих. Dev Biol 344(2): 941-947. 10.1016/j.ydbio.2010.06.010
- McKinney MC, Stark DA, Teddy J, Kulesa PM ( 2011 ). Коммуникация клеток нервного гребня включает обмен цитоплазматическим материалом через клеточные мосты, выявленные с помощью фотоконверсии KikGR. Дев Дин 240(6): 1391-1401. 10.1002/DVD.22612
- Данильчик М., Уильямс М., Браун Э. ( 2013 ). Филоподии, охватывающие бластоцель, на стадии дробления Xenopus laevis: потенциальная роль в распределении и обнаружении морфогенов. Dev Biol 382(1): 70-81. 10.1016/j.ydbio.2013.07.024
- Ван Т., Сюй З., Цзян В., Ма А. ( 2006 ). Межклеточный контакт индуцирует мезенхимальные стволовые клетки к дифференцировке в кардиомиоциты и гладкомышечные клетки. Int J Cardiol 109(1): 74-81. 10.1016/j.ijcard.2005.05.072
 Eur J Cell Biol 94(10): 429-443. 10.1016/j.ejcb.2015.06.010
Eur J Cell Biol 94(10): 429-443. 10.1016/j.ejcb.2015.06.010 Карр Биол 11(12): R470-472. 10.1016/s0960-9822(01)00284-6
Карр Биол 11(12): R470-472. 10.1016/s0960-9822(01)00284-6 Cell Res 20(1): 72-88. 10.1038/кр.2009.112
Cell Res 20(1): 72-88. 10.1038/кр.2009.112 , Пурбху М.А., Шово А., Колер К., Оддос С., Эйссманн П., Бродский Ф.М., Хопкинс С., Онфельт Б., Саттентау К., Дэвис Д.М. ( 2008 ). Мембранные нанотрубки физически соединяют Т-клетки на большие расстояния, представляя собой новый путь передачи ВИЧ-1. Nat Cell Biol 10(2): 211-219. 10.1038/ncb1682
, Пурбху М.А., Шово А., Колер К., Оддос С., Эйссманн П., Бродский Ф.М., Хопкинс С., Онфельт Б., Саттентау К., Дэвис Д.М. ( 2008 ). Мембранные нанотрубки физически соединяют Т-клетки на большие расстояния, представляя собой новый путь передачи ВИЧ-1. Nat Cell Biol 10(2): 211-219. 10.1038/ncb1682 , Буссе Л., Лориа Ф., Чжу С., де Шомон Ф., Пьери Л., Оливо-Марин Дж. К., Мелки Р., Зурзоло С. ( 2016 ). Туннельные нанотрубки распространяют фибриллярный альфа-синуклеин посредством межклеточного переноса лизосом. Embo J 35(19): 2120-2138. 10.15252/прил.201593411
, Буссе Л., Лориа Ф., Чжу С., де Шомон Ф., Пьери Л., Оливо-Марин Дж. К., Мелки Р., Зурзоло С. ( 2016 ). Туннельные нанотрубки распространяют фибриллярный альфа-синуклеин посредством межклеточного переноса лизосом. Embo J 35(19): 2120-2138. 10.15252/прил.201593411 Опен Биол 7(3): 160328. 10.1098/rsob.160328
Опен Биол 7(3): 160328. 10.1098/rsob.160328 , Прадель Г., де Рой Б., Шеффер П., Скебердис В.А., Смит И.Ф., Танвир А. и др.( 2017 ). Туннельные нанотрубки и щелевые соединения — их роль в дальних межклеточных коммуникациях во время развития, здоровья и болезней. Front Mol Neurosci 10: 333. 10.3389/fnmol.2017.00333
, Прадель Г., де Рой Б., Шеффер П., Скебердис В.А., Смит И.Ф., Танвир А. и др.( 2017 ). Туннельные нанотрубки и щелевые соединения — их роль в дальних межклеточных коммуникациях во время развития, здоровья и болезней. Front Mol Neurosci 10: 333. 10.3389/fnmol.2017.00333 Cell Death Differ 22(7): 1181-1191. 10.1038/cdd.2014.211
Cell Death Differ 22(7): 1181-1191. 10.1038/cdd.2014.211 Myo10 является ключевым регулятором образования ТНТ в нейрональных клетках. J Cell Sci 126 (Pt 19): 4424-4435. 10.1242/jcs.129239
Myo10 является ключевым регулятором образования ТНТ в нейрональных клетках. J Cell Sci 126 (Pt 19): 4424-4435. 10.1242/jcs.129239 10.1146/annurev.cellbio.13.1.147
10.1146/annurev.cellbio.13.1.147 Визуализация динамики филоподий в бластоцисте мыши. Dev Biol 265(1): 75-89. 10.1016/j.ydbio.2003.09.012
Визуализация динамики филоподий в бластоцисте мыши. Dev Biol 265(1): 75-89. 10.1016/j.ydbio.2003.09.012 , Вебер С., Шуг А., Раз Э., Шолпп С. ( 2015 ). Транспорт Wnt на основе филоподий во время формирования паттерна ткани позвоночных. Нац Коммуна 6: 5846. 10.1038/ncomms6846
, Вебер С., Шуг А., Раз Э., Шолпп С. ( 2015 ). Транспорт Wnt на основе филоподий во время формирования паттерна ткани позвоночных. Нац Коммуна 6: 5846. 10.1038/ncomms6846 10.1016/j.celrep.2018.07.015
10.1016/j.celrep.2018.07.015 Регуляция формы клеток с помощью Cdc42 опосредуется синергической активностью связывания актина комплексом Eps8-IRSp53. Nat Cell Biol 8: 1337. 10.1038/ncb1502
Регуляция формы клеток с помощью Cdc42 опосредуется синергической активностью связывания актина комплексом Eps8-IRSp53. Nat Cell Biol 8: 1337. 10.1038/ncb1502 Nat Cell Biol 11(12): 1427-1432. 10.1038/ncb1990
Nat Cell Biol 11(12): 1427-1432. 10.1038/ncb1990 Embo J (2019) 38: e101230. 10.15252/прил.2018101230
Embo J (2019) 38: e101230. 10.15252/прил.2018101230 , Манова-Тодорова К., Мур MAS ( 2017 ). Визуализация туннельных мембранных трубок объясняет клеточную коммуникацию в опухолях. Trends Cancer 3(10): 678-685. 10.1016/j.trecan.2017.08.001
, Манова-Тодорова К., Мур MAS ( 2017 ). Визуализация туннельных мембранных трубок объясняет клеточную коммуникацию в опухолях. Trends Cancer 3(10): 678-685. 10.1016/j.trecan.2017.08.001 Коммуникация клеток нервного гребня включает обмен цитоплазматическим материалом через клеточные мосты, выявленные с помощью фотоконверсии KikGR. Дев Дин 240(6): 1391-1401. 10.1002/DVD.22612
Коммуникация клеток нервного гребня включает обмен цитоплазматическим материалом через клеточные мосты, выявленные с помощью фотоконверсии KikGR. Дев Дин 240(6): 1391-1401. 10.1002/DVD.22612Ольга Коренькова является участником Международной программы докторантуры Парижского университета Пастера (PPU).
Анну Пепе поддержали стипендии из Франции Паркинсон. Работа над TNT в лаборатории Chiara Zurzolo поддерживается грантами от Agence Nationale de Recherche (JPND Neutargets: ANR-14-JPCD-0002-01 и ANR-16 CE160019-01 NEURO-TUNN) и Equipe FRM (Fondation Recherche Mé-dicale) 2014 (DEQ20140329557).